Introduction
Inflammation is an immune response that protects host from harmful stimulants such as infection, cellular damage, and tissue injury1–2). The inflammatory response induces activation and recruitment of immune cells, resulting in production of a variety of inflammatory mediators3). In particular, pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α and interleukin (IL)-6 were often observed in the pathogenesis of chronic liver inflammation4). Moreover, the etiology of liver fibrosis, cirrhosis, and hepatocellular carcinoma (HCC) is also related to inflammatory pathology, showing the elevation of TNF-α and IL-65–7).
Hepatic fibrosis, a healing process for liver damage, is the result driven by inflammation responses in liver. As a result of the inflammation process, macrophage-derived transforming growth factor (TGF)-β1 stimulates the activation of hepatic stellate cells (HSCs)8–9). Activated HSCs result in excessive accumulation of the extracellular matrix (ECM) including α-smooth muscle actin (SMA), type I collagen (collagen I), matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs)10–11. Liver fibrosis is a reversible state, but the possibility of recovery is substantially reduced if it progresses to cirrhosis or HCC12–14). Although efficient therapeutic strategies are urgently required, until now there is no standard treatment for ameliorating liver inflammation and fibrosis simultaneouly15–17).
IL-6 and TNF-α are pro-inflammatory cytokines mediating inflammatory responses and are secreted in macrophages and Kupffer cells6),18). IL-10 is a cytokine secreted by macrophage activation and liver injury processes which can intervene pro-inflammatory macrophage function. Collagen I and α-SMA, the most abundant factors of ECM components, are mainly produced in activated stellate cells and are responsible for cell structure, motility and contractility9),19). MMP-2 is also expressed in activated stellate cell, related to HSCs proliferation20) and regulated by TIMPs21). TIMP1 is an inhibitor of activated HSCs-apoptosis and appears to be an important factor in promoting hepatic fibrosis 10),22–23).
Kyungheechunggan-tang (KHCGT)-A is composed of 9 kinds of herbs (Table 1) which was used for the treatment of acute/chronic liver diseases24). In the previous studies, KHCGT-A was demonstrated to show hepato-protective effects by reducing cellular steatosis, reactive oxygen species (ROS) production, and apoptosis24) and anti-fibrotic effects by suppressing collagen I and α-SMA mRNA expression9). The most important components among herbs composed of KHCGT-A are thought to be Artemisia capillaris (AC), Alisma orientale (AO), and Curcuma longa (CL). AC has been represented to exhibit anti-inflammatory25) and anti-fibrotic effects26) and to have the most significant anti-fibrotic effect among the KHCGT-A constitutive agents9). AO has been implicated in the protection against inflammation and fibrosis on non-alcoholic fatty liver disease (NAFLD)27), and CL also has been demonstrated to have anti-inflammatory28) and anti-fibrotic effects9). In addition, hepato-protective agents Crataegus pinnatifida (CP) and Lycium chinense (LC) were also considered to be important, because CP has shown anti-inflammatory potentials in vitro and in vivo against lipopolysaccharide (LPS) treatment29) and LC has been reported to show anti-inflammatory30) and anti-fibrotic effects31) against hepatic disorders. Based on these herbs, KHCGT-B (AC, AO, CL, CP, and LC) and KHCGT-C (AC, AO, and CL), were newly developed.
The aim of this study is to confirm the anti-inflammatory and anti-fibrotic effects of KHCGTs (KHCGT-A, -B, and -C) on RAW 264.7 cells and LX-2 cells. In order to evaluate their anti-inflammatory effects, RAW 264.7 cells and LX-2 cells were induced by LPS followed by KHCGTs treatment and for the purpose of observing the anti-fibrotic effects, KHCGTs were applied to LX-2 cells.
Materials and methods
1. Preparation of herbal extracts
All herbal plants consisting of Kyungheechunggantang (KHCGT) (Table 1) were purchased from Kyung Hee Herb Pharm (Wonju, South Korea), an herbal company operated by Korea Good Manufacturing Practice (KGMP) guideline. KHCGT-A 101 g, KHCGT-B 96 g, and KHCGT-C 75 g were extracted for 3 h with 30 % ethanol of 1000 ml, 1000 ml, and 750 ml, respectively. Each extracted solutions were filtrated, followed by the repeated extraction process once more. Herbal samples were then evaporated, freeze-dried in order. After processing in freeze dryer apparatus, a form of dried extract powders of each herbal decoction was obtained as 33.72 g of KHCGT-A, 25.38 g of KHCGT-B and 14.40 g of KHCGT-C with extraction yield of 33.39 %, 26.44 % and 19.20 %, respectively.
2. Cell culture
2.1. LX-2 cells
LX-2 cells, immortalized human hepatic stellate cells, were kindly provided by Professor Sang Geon Kim (College of Pharmacy, Seoul National University, Seoul, South Korea). LX-2 cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM; Hyclone, MA, USA) supplemented with 10 % fetal bovine serum (FBS; Equitech-bio, Texas, USA) and 1 % antibiotics (Hyclone, MA, USA) in an atmosphere of 5 % CO2 at 37 °C.
3. Lipopolysaccharide (LPS) and herbal treatment
RAW 264.7 cells and LX-2 cells were pretreated with or without three different herbal extracts (KHCGT-A, -B, and -C) for 24 h, and challenged with LPS (Sigma, St Louis, MO, USA) 50 ng/ml and 100 ng/ml treatment for 18 h and 4 h, respectively.
4. Cell viability assay
Cell viability was determined by 3-(4,5- dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. LX-2 cells and RAW 264.7 cells were seeded in 96-well plates at a density of 5103) cells and 1104) cells per well in DMEM media, respectively. After 24 h of stabilization, three kinds of herbal samples were applied with various concentrations (0, 7, 15, 31, 62, 125, 250, 500 μg/ml) and the cells were incubated for another 24 h and 48 h. Subsequently, 100 μl of MTT solution (Promega, Madison, WI, USA) was added for 2 h, and the cells were successively incubated for another 2 h at 37 °C in a humid atmosphere of 5 % CO2. The insoluble formazan crystals were dissolved in 150 μl/well dimethysulfoxide (DMSO; Lonza, Allendale, NJ, USA) and absorbance value was read at 540 nm by a microplate reader (BMG LABTECH, Ortenberg, Germany).
5. ELISA assay for IL-6, TNF-α, and IL-10
RAW 264.7 cells and LX-2 cells were plated in 24 well plates at a density of 2.5104) cells per well in 500 μl of DMEM media containing 10 % FBS and 100 U/ml of antibiotics for 24 h. After overnight acclimation, the cells were pretreated with 125 and 250 μg/ml of herbal medicine samples for another 24 h, and then stimulated with LPS 50 ng/ml for 18 h and 100 ng/ml for 4 h for RAW 264.7 cells and LX-2 cells, respectively. The supernatants from each well were collected and centrifuged at 12000 rpm and 25 ° for 1 min. The mouse ELISA kit (OptEIA mouse IL-6, TNF-α, IL-10 ELISA set and human IL-6 set, BD, CA, USA) was used for the quantitative determination of IL-6, TNF-α, and IL-10 levels contained in the supernatants, according to the manufacturer’s protocol. After treating the stop solution, the quantitative expression amounts of IL-6, TNF-α, and IL-10 were measured at 450 nm by a microplate reader (BMG LABTECH, Ortenberg, Germany).
6. Quantitative real-time PCR analysis
Total cellular RNA was extracted from the LX-2 cells using the Hybrid-R kits (GeneAll, Korea), and isolated RNA was reverse-transcribed using RT dryMIX (Enzynomics, Korea). The purity and quantity of RNA samples were determined by Nanodrop 2000 (Thermo Scientific). Quantitative real-time PCR was performed using CFX96 TouchTM Real-Time PCR Detection System (BIORAD, USA) which was set up with 40 cycles.
Collagen1A1 and α-SMA: First cycle at 95 °C for 3 min, 40 cycles at 95 °C for 15 sec, 59 °C for 10 sec, 72 °C for 40 sec, and last cycle at 95 °C for 10 sec
TIMP1 and MMP-2: First cycle at 95 °C for 3 min, 40 cycles at 95 °C for 15 sec, 61.4 °C for 10 sec, 72 °C for 40 sec, and last cycle at 95 °C for 10 sec
The primers used for complementary DNA amplication of α-SMA, Collagen1A1, TIMP1, MMP-2, and β-actin genes are presented in Table 2. The mRNA expression results of each gene were normalized with β-actin mRNA level.
Results
1. Effects of KHCGT on cell viability
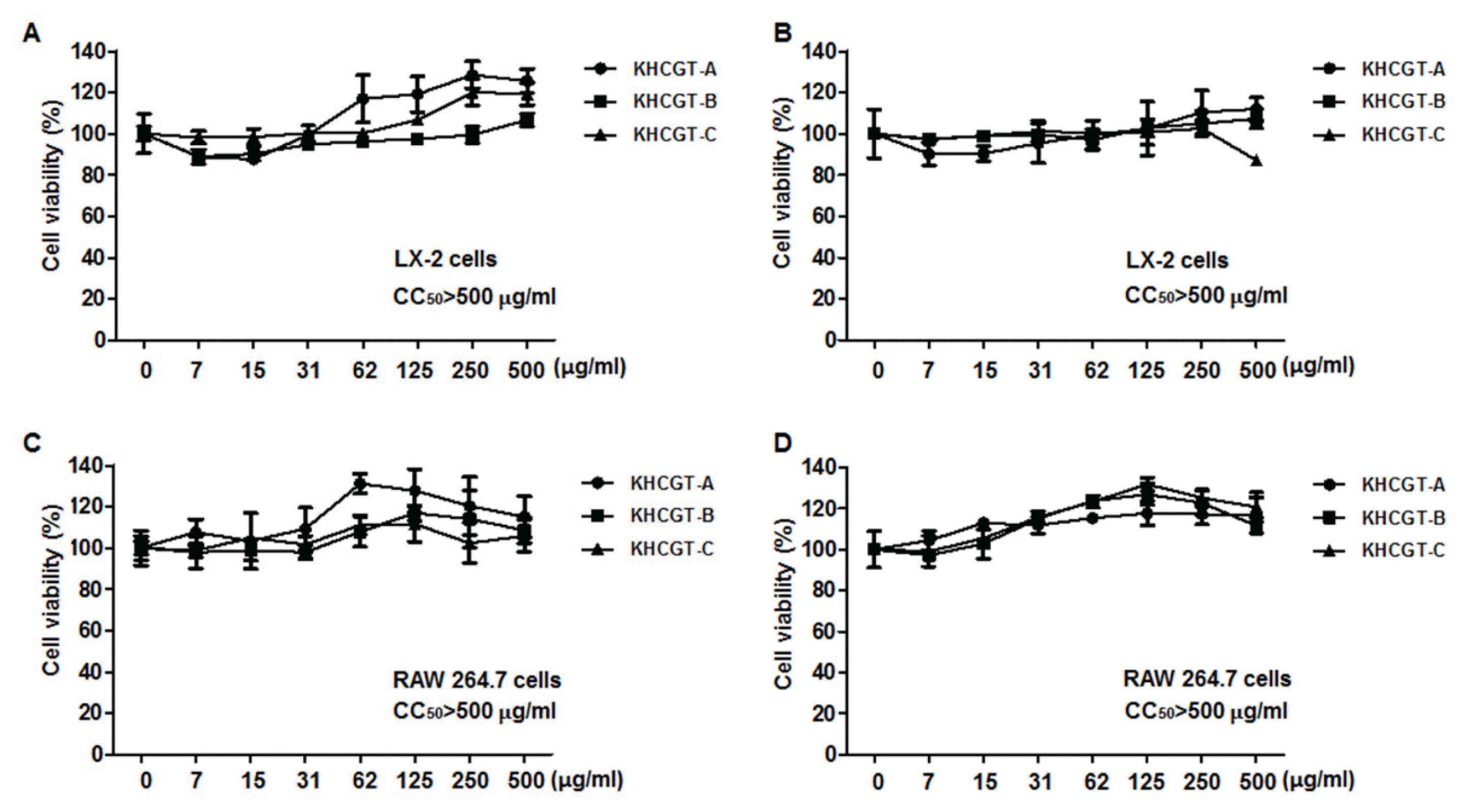
To clarify safeness of KHCGT on LX-2 cells and RAW 264.7 cells, MTT assay on both cells was performed with all three kinds of KHCGTs (KHCGT-A, -B and -C) for 24 h and 48 h, respectively. Treatment on LX-2 cells of KHCGTs with different concentrations (0, 7, 15, 31, 62, 125, 250, 500 μg/ml) exhibited no significant decrease in cell viability in all three types of KHCGTs after overnight and 2 days (Figs. 1A and 1B), suggesting that herbal samples affect no direct toxic changes on HSCs. Similarly, all sorts of KHCGTs at the same concentrations did not cause cytotoxicity in RAW 264.7 cells for 24 h and 48 h (Figs. 1C and 1D). The 50 % cytotoxic concentration (CC50) values were estimated more than 500 μg/ml (Fig. 1). These data indicate that three KHCGT formula samples have no cytotoxicity on both LX-2 cells and RAW 264.7 cells.
2. Effects of KHCGT on inflammatory mediators in LPS-stimulated RAW 264.7 cells
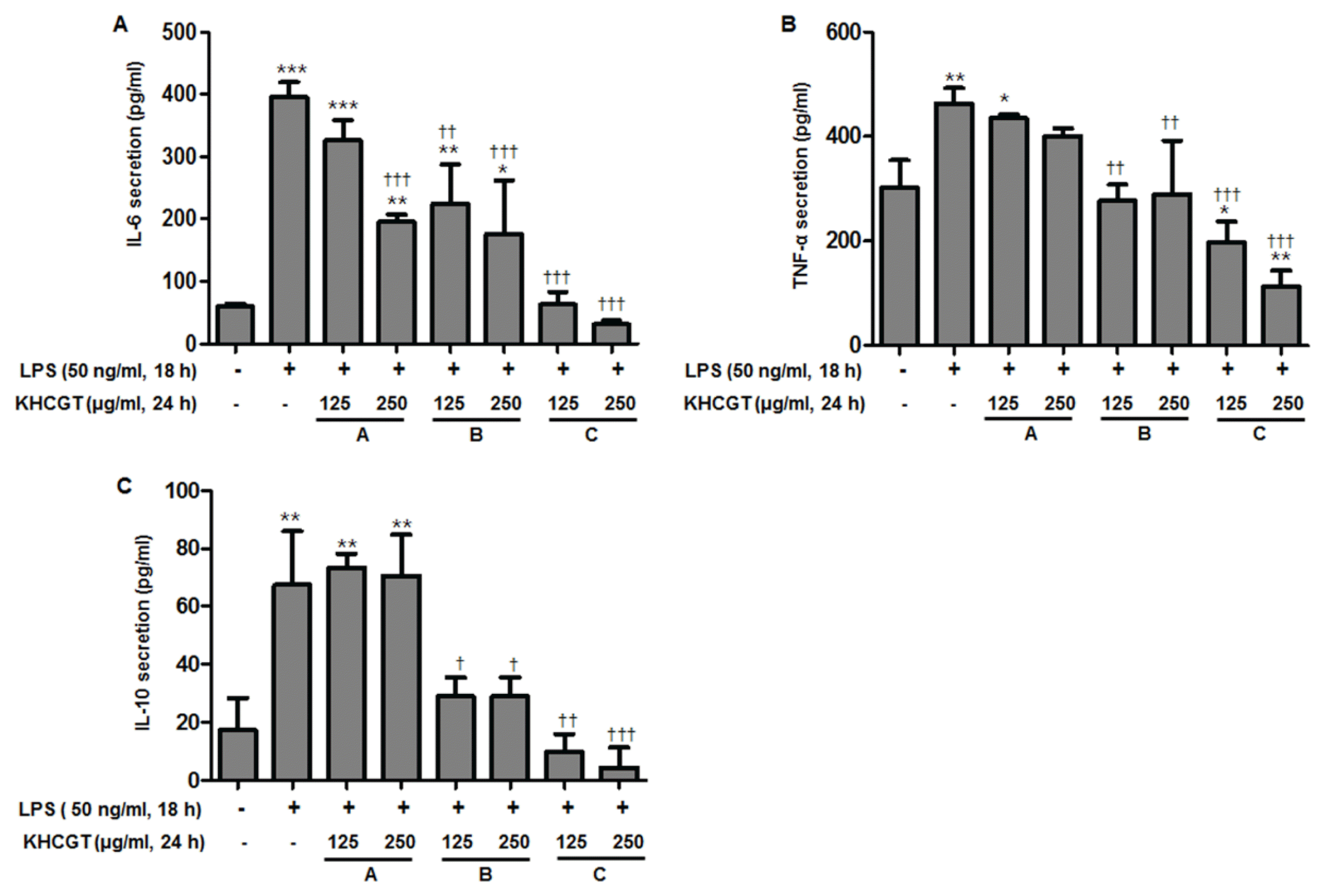
To determine anti-inflammatory effects of KHCGT extracts, the productions of IL-6, TNF-α, and IL-10 in LPS-stimulated RAW 264.7 cells were evaluated by ELISA. In response to LPS 50 ng/ml treatment, RAW 264.7 cells expressed increased cytokine release and these pro-inflammatory cytokines have been reported to contribute to a variety of inflammation-related diseases. IL-6 level was significantly reduced after 24 h pre-treatment with KHCGT-A, -B, and -C in a concentration-dependent manner. Particularly, KHCGT-C treatment showed the most potent inhibitory effect among three herbal formulas against IL-6 secretion, exhibiting low detection of IL-6 expression similar to the control group without LPS treatment (Fig. 2A). As shown in Figs. 2B and 2C, KHCGT-B and -C showed significant suppression on TNF-α level compared to LPS-treated RAW 264.7 cells. In accordance with the effects of KHCGT-C on IL-6 production, 250 μg/ml of KHCGT-C revealed the most marked inhibitory effects on TNF-α and IL-10 levels among three samples in a dose-dependent manner (p < 0.001).
3. Effects of KHCGT on IL-6 production in LPS-induced LX-2 cells
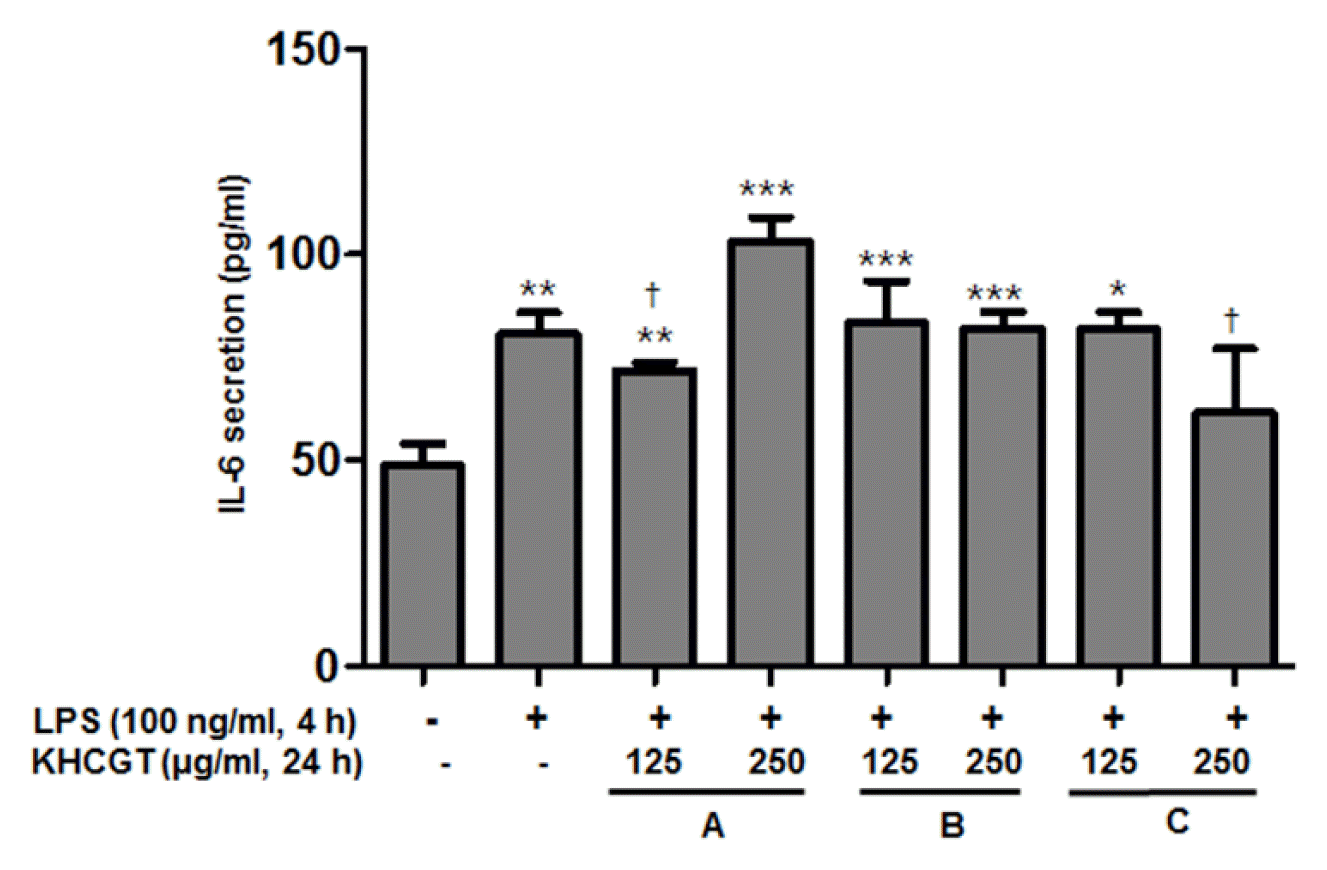
It is known that LPS-induced upregulation of IL-6 is involved in HSCs activation, a critical initial signal in the pathogenesis of liver fibrosis8). To examine whether KHCGT-A, -B, and -C inhibit IL-6 levels produced in response to LPS stimulants on LX-2 cells, ELISA was performed with the supernatants of these formula-treated cells. LPS treatment of 100 ng/ml for 4 h on LX-2 cells triggered significant secretion of IL-6 (p < 0.01). As shown in Fig. 3, 125 μg/ml of KHCGT-A and 250 μg/ml of KHCGT-C significantly inhibited the increased level of IL-6 (p < 0.05).
4. Effect of KHCGT on fibrosis-related genes in LX-2 cells
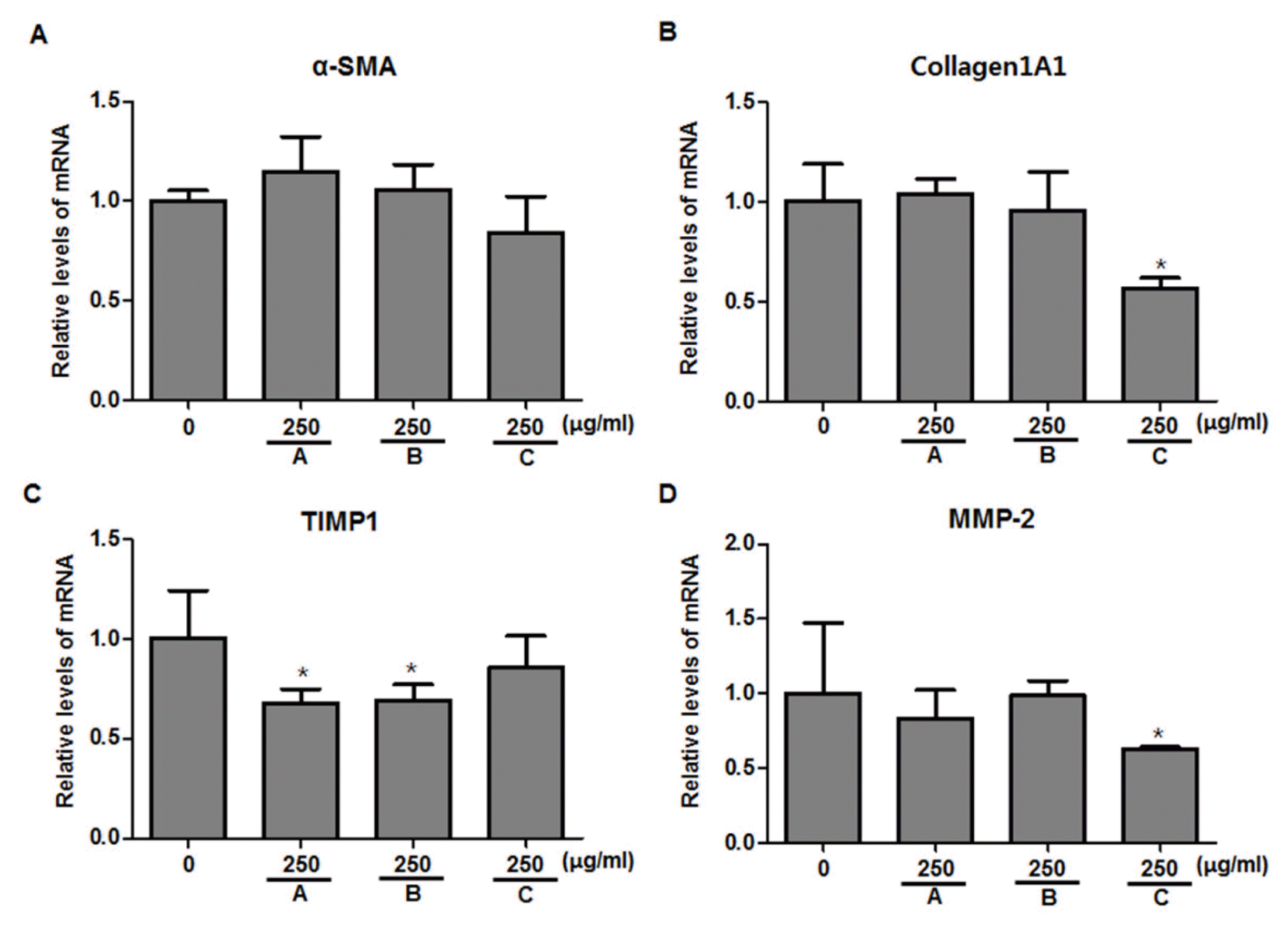
Real-time PCR was performed to evaluate the inhibitory effects of KHCGT-A, -B, and -C on the mRNA levels of α-SMA, collagen1A1, TIMP1, and MMP-2 in LX-2 cells. Each herbal extract was applied to cells at 250 μg/ml which concentration was shown to have significant inhibitory effects on pro-inflammatory mediators (Fig. 2). As illustrated in Fig. 4, KHCGT-C 250 μg/ml significantly inhibited the mRNA expressions of collagen1A1 and MMP-2 (p < 0.05, Figs. 4B and 4D), but there was no significant changes on α-SMA and TIMP1 mRNA levels (Figs. 4A and 4C). Regarding TIMP1 mRNA levels, KHCGT-A and -B showed significant reduction on relative mRNA expression compared to the control group (p < 0.05, Fig. 4C).
Discussion and conclusion
Inflammation and fibrosis play crucial roles in determining liver damage in a wide range of chronic liver diseases, including nonalcoholic fatty liver diseases (NAFLD), viral hepatitis, and alcoholic liver diseases32). Inflammation-driven pathological process in liver activates quiescent HSCs which leads to accumulation of collagen I and α-SMA11). Inversely, stimulated HSCs are often found to produce pro-inflammatory cytokines that keep themselves activated or contribute to the progression to severe fibrosis33). Overall, targeting both inflammation and fibrosis might be of special difficulty but potent therapeutic strategy for treating chronic liver diseases, and numerous herbal medicines such as Saururus chinensis, Salvia miltiorrhiza, and Scutellaria baicalensis have been shown to have inhibitory effects on both inflammation and fibrosis34).
KHCGT-A is a Korean herbal formula composed of 9 medicinal herbs. The Monarch and Minister drug of this decoction is AC and AO, respectively. Besides these two main drugs, KHCGT consists of Atractylodes japonica (AJ), Salvia miltiorrhiza (SM), CL, CP, LC, Poncirus trifoliata (PT) and Glycyrrhiza uralensis (GU). In a previous study, 100 μg/ml water extract of KHCGT-A inhibited steatosis and ROS production by regulating apoptosis in palmitate-induced HepG2 cell models24). In addition, KHCGT-A showed anti-fibrotic effects against activated LX-2 cells and dimethylnitrosamine-induced rat liver by downregulating mRNA levels of collagen I and α-SMA9). KHCGT-A might exert anti-inflammatory activities because AC and AO, main drugs of KHCGT-A, have been reported to regulate nitric oxide, NF-κB, COX-2, prostaglandin E2, and TNF-α25),35–36). However, none of the previous studies demonstrated anti-inflammatory effects of KHCGT-A. Furthermore, there is no precedent study that evaluates its effects on the levels of TIMP-1 and MMP-2, important fibrotic mediators.
To apply KHCGT-A for a wide range of liver diseases related to inflammation and fibrosis, this study is aimed at investigating whether KHCGT-A inhibits the production of IL-6, TNF-α, and IL-10 in LPS-stimulated RAW 264.7 cells and α-SMA, collagen1A1, TIMP1, and MMP-2 in LX-2 cells for the first time. In addition, its actions on enhanced IL-6 synthesis are analyzed in LPS-induced LX-2 cells. Furthermore, these experiments are repeated in the samples treated with KHCGT-B and -C, new formulas condensed from KHCGT-A.
IL-6 and TNF-α are crucial pro-inflammatory mediators involved in liver inflammation such as NAFLD and viral hepatitis37) as well as chronic liver fibrosis. Especially, IL-6 stimulated collagen synthesis in rats38) and higher hepatic TNF-α was observed in more severe liver fibrosis39). On the other hand, IL-10 inflammatory cytokines are recently shown to be related to tissue remodeling, protecting from liver fibrogenesis. Collagen1A1 and α-SMA are predominant markers in fibrosis, mainly expressed in the transition from quiescent HSCs into myofibroblasts11). Early increase of MMP-2 promotes fibrosis and TIMP1 activates anti-apoptosis of HSCs, thus increasing fibrogenic cell survival40–41).
As shown in Fig. 2, KHCGT-A of 250 μg/ml significantly inhibited IL-6 enhanced by LPS in RAW 264.7 cells (p <0.001), while the level of IL-10 secretion was maintained similar to LPS-induced group after treating KHCGT-A (Fig. 3). Anti-fibrotic activities of KHCGT-A through the maintenance of IL-10 seem to be associated with significant reduction of TIMP1 in LX-2 cells (p <0.05, Fig. 4). Both KHCGT-B and -C significantly suppressed IL-6, TNF-α, and IL-10 levels activated in LPS-induced RAW 264.7 cells. Interestingly, KHCGT-C composed of only 3 herbs led to the most drastic reduction in the level of pro-inflammatory factors in RAW 264.7 cells and LX-2 cells, both stimulated by LPS treatment. However, KHCGT-C significantly suppressed IL-10 levels, anti-fibrotic mediators, as well as collagen1A1 and MMP-2 mRNA expression. Probably, there are various steps of complicated inflammatory and fibrogenic pathological process, and each formula can intervene different steps.
In conclusion, this study demonstrates that KHCGT shows an inhibitory effect on the expression of pro-inflammatory cytokines such as IL-6, TNF- α, and IL-10 in LPS-stimulated RAW 264.7 cells as well as fibrosis-related genes in LX-2 cells. Among three formulas, KHCGT-C showed the most outstanding efficacy particularly in suppressing inflammatory mediators in LPS-induced RAW 264.7 and LX-2 cells and the expression of collagen1A1 and MMP-2 mRNA in LX-2 cells. Its pharmacological properties might contribute to the development of anti-inflammatory, hepato-protective, and anti-fibrotic agents. However, the molecular mechanism responsible for suppressing inflammatory mediators by KHCGT treatment has not been yet elucidated. Therefore, further investigations are required to clarify molecular signaling pathway of KHCGT related to anti-inflammatory and anti-fibrotic effects in various pathological models.