Introduction
The inflammatory response is a type of immune response, and is mainly regulated by macrophages as one of the defense mechanisms to normalize the damaged function of cells1). In particular, microglia, the macrophage in the brain, is involved in the inflammatory response of the brain and protect the brain nerve cells and regenerate tissues2). In addition, it is known that the microglia could maintain normal brain functions, including the overall structure, the neurotransmitters, and the neuronal synapses3). However, upon the microglia is over-activated, microglia produces a large amount of pro-inflammatory cytokines and chemokines, which lead to demyelination, synaptic remodeling, and neuronal death, and ultimately result in the neurodegenerative disease such as Alzheimer’s dementia and Parkinson’s diseases4). Thus, many researches on the regulation of microglia are receiving a lot of attentions as it can contribute to the treatment of neurological diseases. Thus, there is a need to study the new medicines or therapeutics with anti-inflammatory and anti-oxidant effects that can control microglia activity.
Kyungok-go (KOG) is the first prescription recorded in Hong’s Collected Empirical Prescriptions (Hong Shi Ji Yan Fang, 洪氏集驗方) and is a solid preparation made from Rehmanniae Radix (生地黃), Ginseng Radix(人蔘), Poria Sclerotium (茯苓), Mel (蜂蜜)5). KOG has been used traditionally to strengthen the gastrointestinal (脾 臟) and renal (腎臟) system, and to rejuvenate the yin (陰) and yang (陽) in Korean medicine. Recently, it has been reported that KOG could inhibit the inflammatory responses, cranial nerve damages, polycystic ovary syndrome, sperm damage and lung damage through NRF2-Keap1 activity6–9). Although many reports have focused on the beneficial and anti-inflammatory activity of KOG on the various diseases, there have been no reports of studies on the regulation of microglia yet.
Herein, in this study, we investigated whether KOG has the anti-inflammatory, and the neuro-protective activity. To archive it, we treated the lipopolysaccharide (LPS) in BV2 cells, the mouse microglia cell line, and then determined the nitrite, pro-inflammatory cytokines, and regulating mechanisms to investigate the anti-inflammatory activity of KOG.
Materials and Methods
1. KOG preparation
The KOG used in the experiment was purchased from Wonkwang University Korean Medicine Hospital (Iksan, Korea). KOG (100 g) was well diluted with water (1L), and it was frozen at −80°C. After that, the extract was freeze-dried to produce a powder with a yield of 26.9 g. The resulting powder was dissolved in distilled water and filtered. Filtrates were stored at 4°C until use.
2. MTT assay
BV2 microglia cell line was obtained from the Korean Cell Line Bank (Seoul, Korea). The cells were seeded at a density of 2 × 105/mL in RPMI-1640 medium Gibco (Thermo Fisher Scientific, MA, USA), and then treated with KOG. After incubating for 24 h, 3-(4,5-dimethylthiazol-2-yl)- 2,5-diphenyl-2H-tetrazolium bromide (MTT, Sigma, MO, USA) solution was added for 30 min. After removing the supernatant, dimethyl sulfoxide (DMSO, Sigma, MO, USA) was added to dissolve formazan, and the solution was loaded into a 96-well plate and absorbance was measured at 540 nm.
3. Measurement of Nitrite Concentration
BV2 cells were cultured at a density of 2×105/mL in RPMI-1640 medium, and then treated with KOG for 1 h. After the stimulation with 1 μg/mL of LPS (Sigma, MO, USA) for 24 h, the cell supernatant was loaded into a 96-well plate. After adding 100 μL of griess reagent (Sigma, MO, USA), absorbance was measured at 540 nm.
4. RNA isolation and quantitative polymerase reaction (Quantitative RT-PCR)
BV2 cells were cultured at a density of 1×106/mL in RPMI-1640 medium, and then treated with KOG for 1 h. After the stimulation with LPS for 6 h, the cells were washed with phosphate buffered saline (PBS), and 1 mL of Easy Blue (Intron Biotechnology, Seongnam, Korea) was added to lyses the cells to extract RNA. Reverse transcription of RNA to cDNA was performed using an ABI cDNA synthesis kit Applied Biosystems, MA, USA). Subsequently, the cDNA was amplified using the SYBR Premix kit (Applied Biosystems, MA, USA) using a StepOne Plus Real-Time PCR system (Applied Biosystems, MA, USA). 30 s at 95°C, 40 cycles at 95°C for 5 s and 60°C for 60 s each, dissociation for 15 s at 95°C and 30 s at 60 °C, and then 15 s at 95°C on ABI Step one Plus. StepOne software (Applied Biosystems, MA, USA) was used for data analysis. Relative gene expression (target gene expression normalized to that of the endogenous control gene) was calculated using the comparative Ct method (2−ΔΔCt). The analysis was independently conducted three times. Forward(f) and reverse(r) primers synthesized by Cosmogenetec (Daejeon, Korea). The primers used are shown in Table 1.
5. Western blot analysis
BV2 cells were seeded at a density of 5 x 106/mL in RPMI-1640 medium and starved with serum free media for 12 h. Subsequently, the cells were pre-treated with KOG for 1 h and stimulated with LPS. After 0, 15, 30, or 60 min, the cells were captured after washing 3 times with cold PBS. Then the cells were lysed with RIPA lysis buffer (Sigma, MO, USA) to dissolve the protein. After mixing with sampling buffer (4X), the sample was electrophoresed on 10% SDS-PAGE, transferred to a membrane, and blocked with 5% skim milk for 2 h. The protein expressions were confirmed with the ECL detection solution (Amersham, UK). Anti-phospho -extracellular signal regulated kinase (ERK) 1/2, anti-phospho-p38, anti-phospho-c-Jun N-terminal kinase (JNK) antibodies used in the experiment were manufactured by Cell Signaling (Danvers, MA, USA). Anti-Iκ-Bα, Anti-β-actin, Anti-ERK, Anti-JNK, and Anti-p38 were purchased from Santa Cruz biotechnology (Dallas, TX, USA).
Results
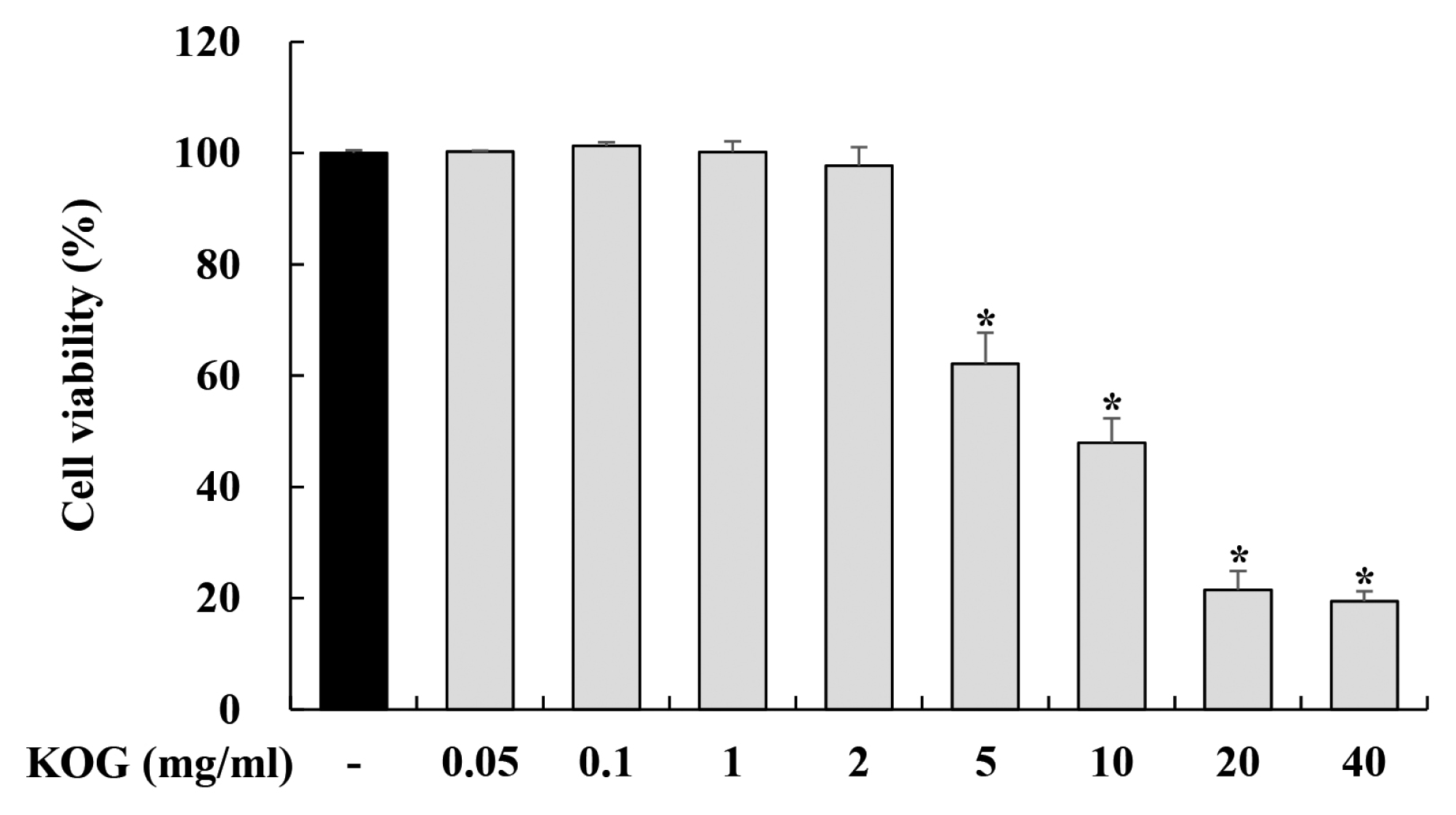
1. Cytotoxicity of KOG in BV-2 microglia
In order to determine the concentration range of KOG in BV2 microglia, we performed the MTT assay to evaluate the cytotoxicity of KOG. BV2 cells were treated with KOG at a concentration of 0.05–40 mg/mL for 24 h. As shown in Fig 1, up to a dose of 2 mg/mL, KOG did not affect the cell viability. (Fig. 1). Therefore, we chose the concentrations ranges of KOG from 0 to 2 mg/mL for further experiments.
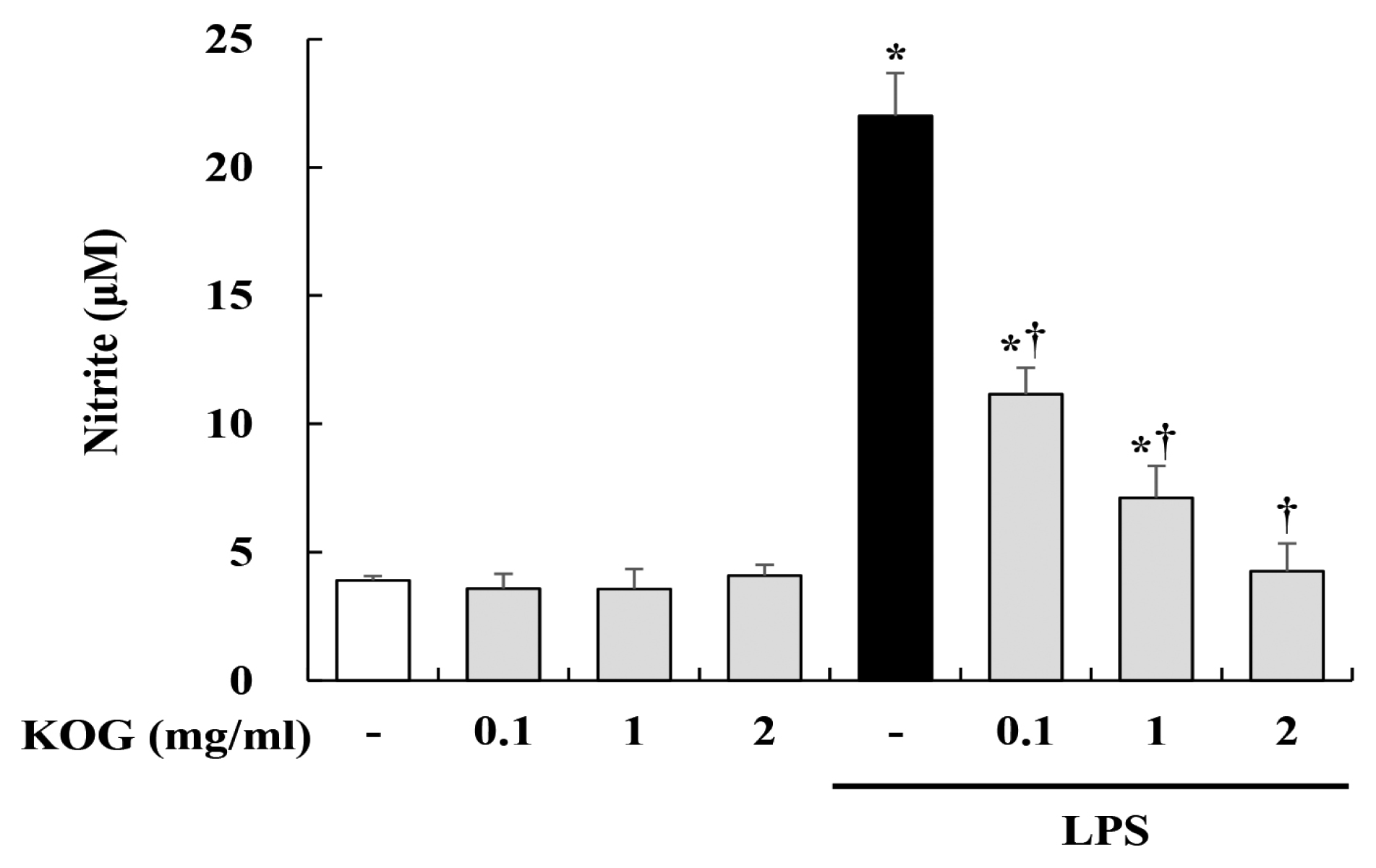
2. Inhibitory effect of KOG on nitrite production in LPS-stimulated BV2 microglia
In order to investigate the anti-inflammatory effect of KOG, we measured the nitrite production by Griess assay. BV2 microglia were pretreated with KOG at the concentrations of 0.1, 1, and 2 mg/mL for 1 h, and then stimulated with LPS for 24 h. As shown in Fig 2, our data showed that LPS treatment markedly induced the nitrite production in BV2 cells. However, the increase of nitrite by LPS was significantly decreased by KOG in concentration-dependent manner (Fig. 2).
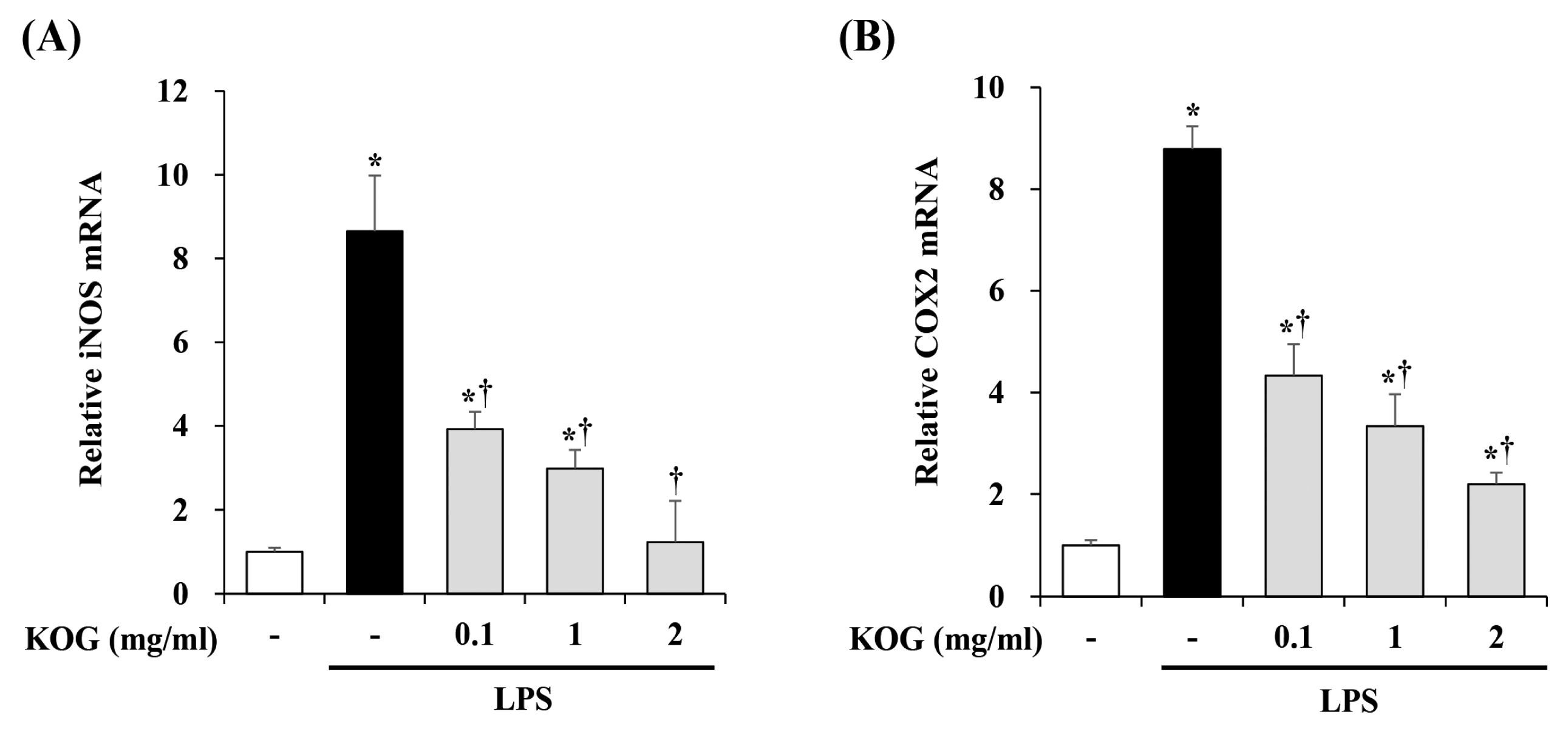
3. Inhibitory effect of KOG on iNOS and COX-2 expression in LPS-stimulated BV2 microglia
To investigate the inflammatory mediators such as inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2, we performed the Quantitative RT-PCR. BV2 microglia were pretreated with KOG at the concentrations of 0.1, 1, and 2 mg/mL for 1 h, and then stimulated with LPS for 6 h. As shown in Fig 3, our data showed that LPS treatment markedly increased the mRNA levels of iNOS and COX-2 in BV2 cells. However, the mRNA levels of both iNOS and COX-2 inhibited by KOG in concentration-dependent manner (Fig. 3).
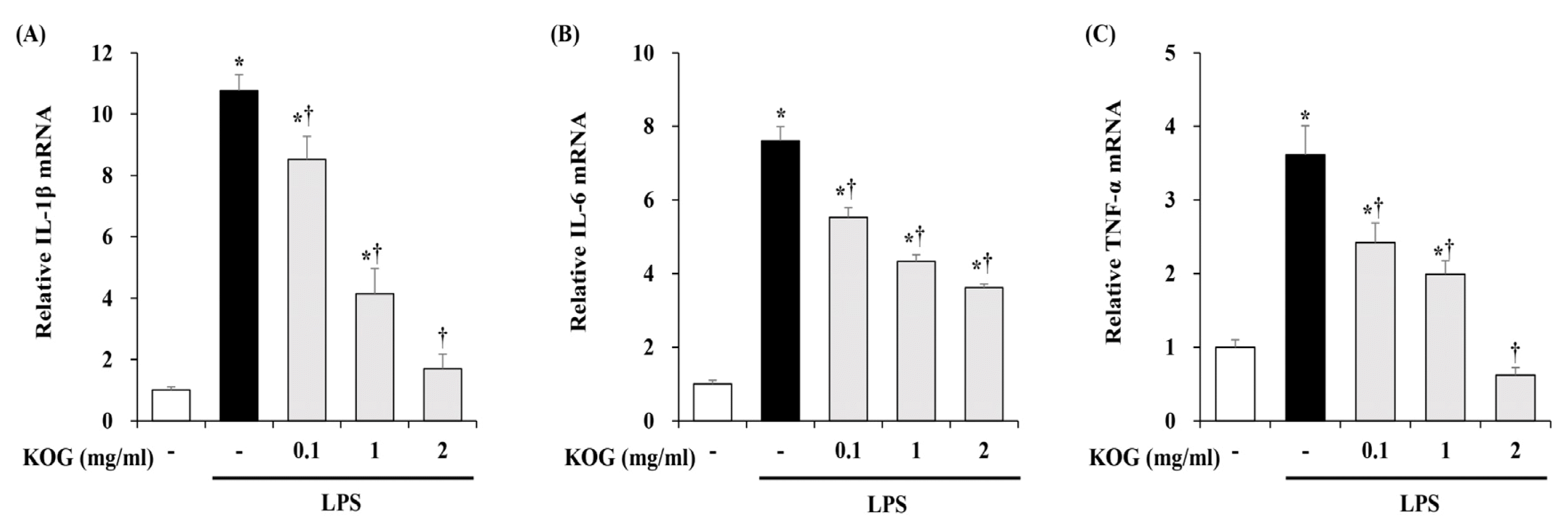
4. Inhibitory effect of KOG on IL-1β, IL-6, and TNF-α expression in LPS-stimulated BV2 microglia
To investigate the pro-inflammatory cytokines such interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α, we performed the Quantitative RT-PCR. BV2 microglia were pretreated with KOG at the concentrations of 0.1, 1, and 2 mg/mL for 1 h, and then stimulated with LPS for 6 h. As shown in Fig 3, our data showed that LPS treatment markedly increased the mRNA levels of IL-1β, IL-6, and TNF-α in BV2 cells. However, the mRNA levels of IL-1β, IL-6, and TNF-α inhibited by KOG in concentration -dependent manner (Fig. 4).
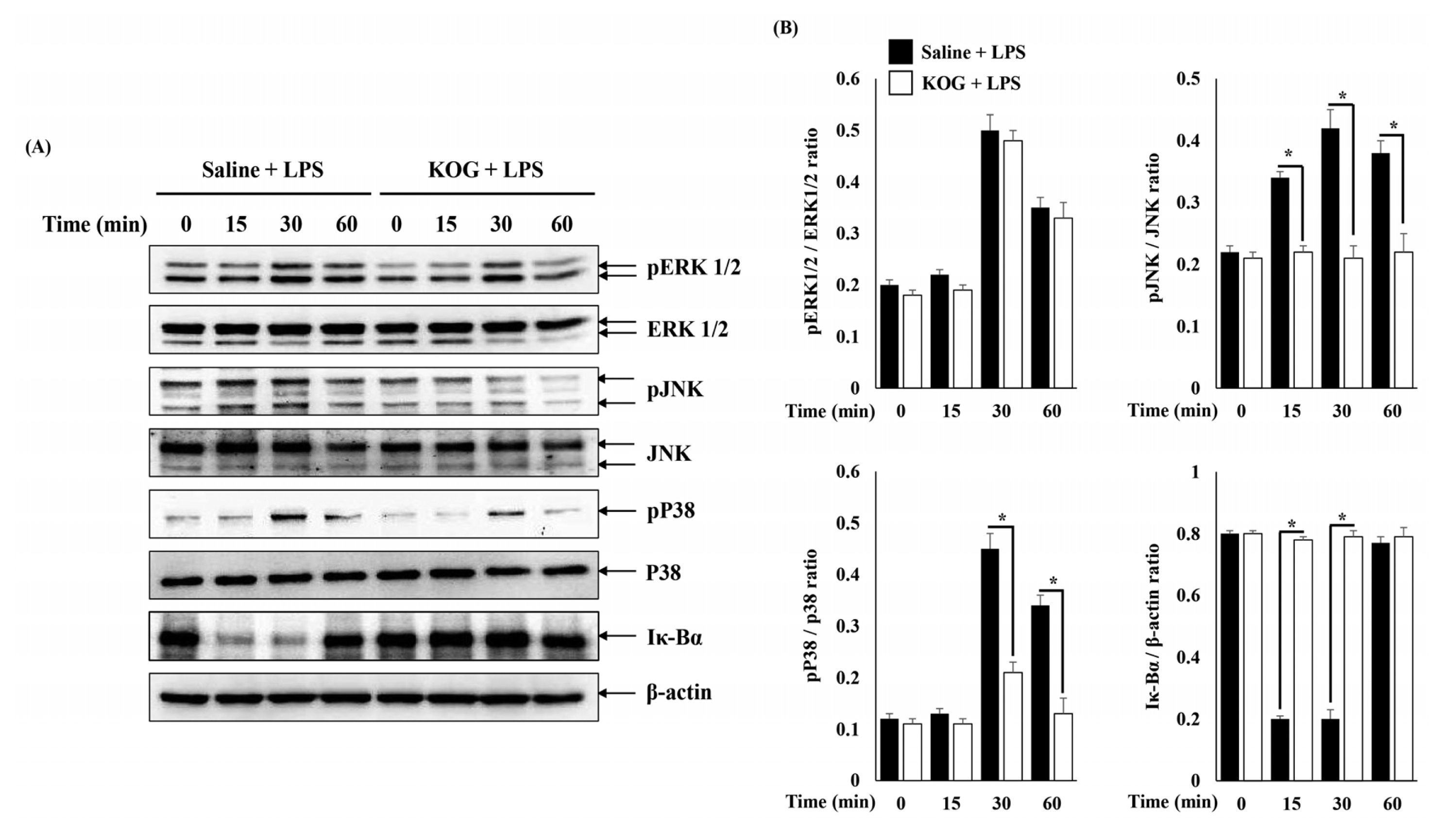
5. Regulatory mechanisms of KOG in LPS-stimulated BV2 microglia
To investigate the regulatory mechanisms of KOG, we examined the activation of mitogen-activated protein kinases (MAPKs; ERK, JNK, p38) and nuclear factor (NF)-κB signaling pathways by western blot. BV2 microglia were pretreated with KOG at the concentrations of 2 mg/mL for 1 h, and then stimulated with LPS for indicated times. As shown in Fig. 5, phosphorylation of ERK, JNK, and p38 in LPS-treated BV2 microglia peaked at 30 min and then decreased slightly at 60 min. In the case of Iκ-Bα, degradation was increased over 15 and 30 min, and degradation was recovered at 60 min. However, the phosphorylation of JNK and p38 and degradation of Iκ-Bα were suppressed by KOG, meanwhile the phosphorylation of ERK1/2 was not altered (Fig. 5).
Discussion
Neurodegenerative diseases such as Alzheimer’s disease, Parkinson’s disease, cerebellar atrophy and Huntington’s disease are associated with neuronal cell death and dysfunction, and are gradually induced by the impairment of brain function10). Although we can’t match the same diseases with the neurodegenerative diseases in Korean medicine, we can predict the degenerative neurological disease from the main symptom, such as tremor (震顫), rach (痓), spasm (痙), scrofula (瘈瘲), convulsion (攣), stroke (中風), and so on11). The main causes of these symptoms are wind (風), fire (火), phlegm (痰), stasis (瘀), and empty (虛), and clinical types can be classified into yin deficiency of liver and kidney (肝腎陰虛), deficiency of energy and blood (氣血 兩虛), phlegm-heat internal damping (痰熱內阻), and stagnation of energy and blood (氣滯血瘀)12). Therefore, we could hypothesis that the main symptoms of degenerative neurological diseases caused by deficiency and fatigue (虛勞) such as yin deficiency of liver and kidney (肝腎陰虛), and deficiency of energy and blood (氣血兩虛) would be treated by KOG, which can improve deficiency and fatigue (虛勞).
KOG(瓊玉膏) is a representative herbal medicine prescription with the compensatory activity of the deficiency (補虛損), and is introduced as “KOG could help to fulfill the body spirit and essences, nourish the pith, regulate the energy, help to maintain the homeostasis, rejuvenates the old, and make up for all deficiencies and diseases (塡精補 髓 調眞養性 返老還童 補百損 除百病)”, “KOG could help to improve the mental and body health, make the white hair to black, make the missing teeth come out again, walk like horse (萬 神俱足 五藏盈溢 髮白復黑 齒落更生 行如奔馬), and “If you intake the KOG several times a day, you feel never hungry and thirsty all day long. The beneficial effects of KOG is indescribable (日進數服 終日不飢渴 功效不可盡述)” in Donguibogam (東醫寶鑑)13). Thus, we could predict that KOG could enhance the yin, thereby nourish the respiratory system (滋陰潤肺), and enhance energy, thereby invigorates the gastrointestinal system (益 氣補脾) and hypothesis that the immunodeficiency and inflammation caused by exhaustion or chronic fatigue could be cured by KOG which could fulfill the yin (陰) and energy (氣). Moreover, among the constituents of KOG, Ginseng Radix (人蔘) which contains ginsenosides, and Rehmanniae Radix (生地黃) are reported to have a memory improvement effect and neuroprotective activity in various animal models of dementia14–16). Also, KOG is reported to have a protective effects in animal models of memory loss induced by scopolamine and forebrain ischemia17,18). As described above, it is predicted that KOG will be effective for degenerative neurological diseases based on the Korean traditional and latest scientific experimental literature, but specific studies on its efficacy and mechanism are still insufficient. Therefore, in this study, we tried to verify the neuroprotection efficacy and mechanism of KOG using microglia.
Although the cause of neurodegenerative diseases is not clear, it has recently received a lot of attention that the damages to nerve cells by the inflammatory response of microglia, which is responsible for immunity within the central nervous system, is the one of various causes in neurodegenerative diseases10). When the inflammatory response of microglia is excessive and not under control, the secretion of inflammatory mediators such as iNOS and COX-2 and pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α, increases, which leads to neurological diseases pathological condition19). Thus, the regulation of these inflammatory mediators and pro-inflammatory cytokines could by a promising method for treating neurological diseases. Therefore, in this study, we investigated the potentials of KOG in LPS-induced micro inflammation model, which mimics the brain inflammation and neurological diseases.
LPS has been widely used as an inflammagen that causes inflammation in the brain20). LPS plays a central role as a pathogen in the inflammatory response and stimulates the production of inflammatory mediators such as nitrite, prostaglandin E2 (PGE2) and leukotriene, and activates the signaling pathways of these inflammatory mediators20). Nitric oxide (NO) produced by the oxidation of nitrite plays an important role in the regulation of physiological synthesis, and is mainly regulated through iNOS during inflammation21). Upon the iNOS level is increased against pathogens, a large amount of NO would be generated to attenuate the local inflammatory response by suppressing the bacterial invasion, and inhibiting T-cell proliferation. However, when the inflammation is not under control, the excessive over-expression of iNOS leads to over-generation of NO, which ultimately results in cellular injury and inflammation22,23). Like iNOS, COX-2 is an inducible substance whose expression is increased by external stimuli such as LPS, cytokines, and chemokines, and its excessive production could cause the inflammation24). In this study, we evaluated the anti-inflammatory activity of KOG in LPS-induced BV2 microglia, the cellular brain inflammation model and determined the nitrite, iNOS and COX-2 expression as a marker of inflammation. As a result, it was found that the LPS-induced nitrite production and the increased expression of iNOS and COX-2 in microglia were significantly inhibited by the treatment with KOG (Fig. 2, 3), which means KOG could attenuate the production of inflammatory mediators.
When inflammatory cells are activated, not only NO but also pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α are over-secreted, which can exacerbate the inflammatory response25). Therefore, in this study, we evaluated the inhibitory effect of KOG on the expression of pro -inflammatory cytokines IL-1β, IL-6, and TNF-α as an additional inflammation marker. As a result, we could find that the elevated expression of IL-1 β, IL-6, and TNF-α induced by LPS in microglia was significantly inhibited by KOG in a concentration-dependent manner (Fig. 4), which suggests KOG could attenuate the production of pro-inflammatory cytokines against LPS.
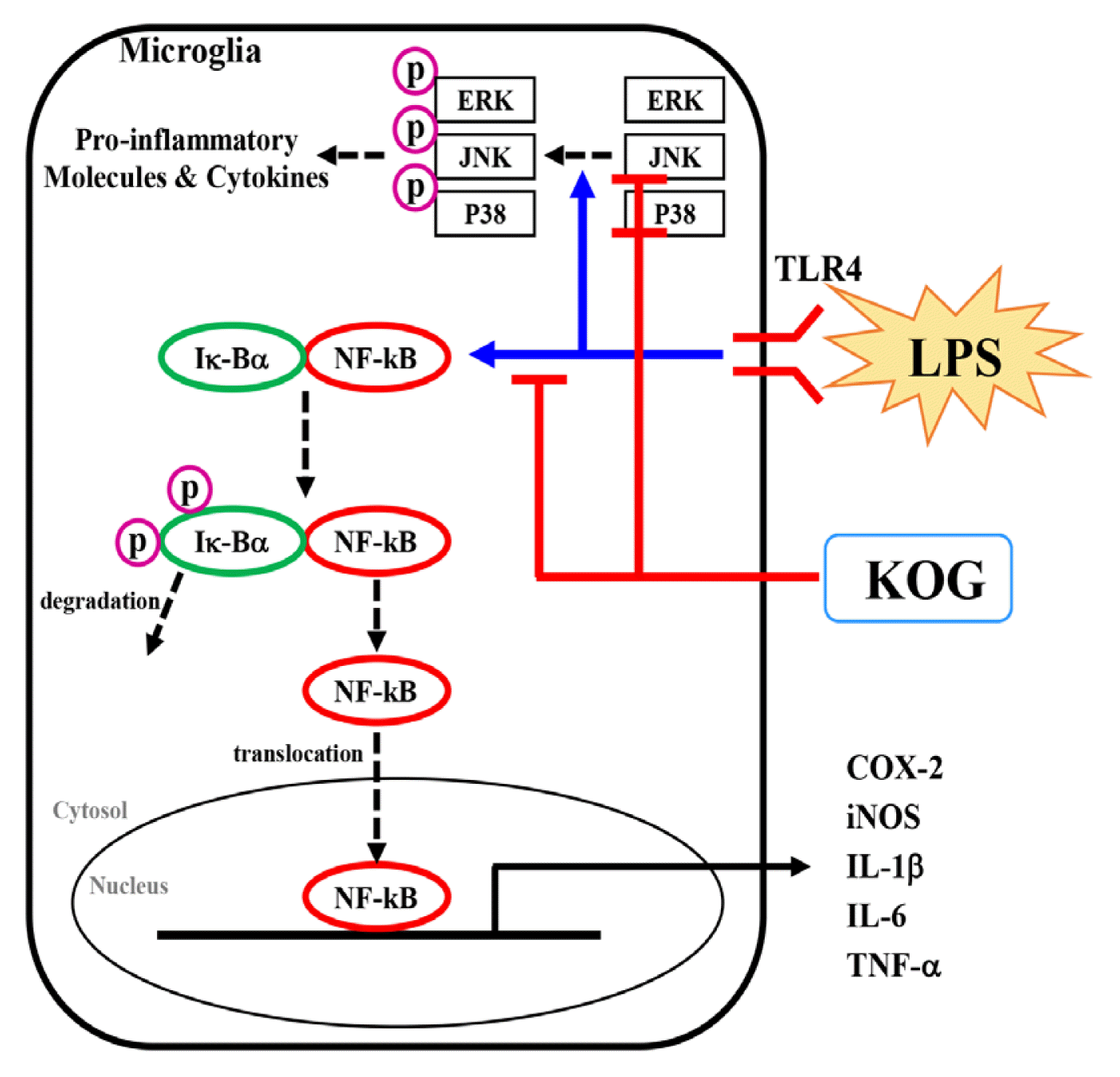
The mitogen-activated protein kinase (MAPK) pathway is known to play an important role in apoptosis, cell differentiation, cell growth, cell proliferation and immune responses26). The three major proteins involved in the MAPK pathway are extracellular signal regulated kinases 1 and 2 (ERK1/2), c-Jun N-terminal kinases (JNKs), and p38 MAPK27). In addition, nuclear factor kappa B (NF-κB) is an essential transcription factor for the secretion of inflammatory mediators. The p65 and p50 proteins, which are components of NF-κ B, generally exist with a bound to the inhibitor kappa-Bα (Iκ-Bα) in the cytoplasm, and then p65 and p50 are separated from Iκ-Bα due to the cellular stimualtion such as inflammation28). Then, the isolated Iκ-Bα become phosphorylated and total amount of Iκ-Bα is degraded28). In addition, the separated p65 and p50 move into the nucleus and bind to DNA, thereby activating the transcription of inflammatory mediators and cytokines29). Accordingly, in order to investigate the anti-inflammatory mechanisms of KOG, we evaluated the activation of MAPKs and NF-κB in LPS-induced BV2 microglia. As a result, the phosphorylation of ERK, JNK, p38 and degradation of Iκ-Bα was significantly increased in the LPS-only treatment group. However, JNK, and p38 phosphorylation and Iκ-Bα degradation were significantly inhibited in the group treated with KOG (Fig. 5). These results could suggest that KOG exhibited an anti-inflammatory effect through the inhibition of JNK, p38 and NF-κB signaling pathways in LPS-induced BV2 microglia.
Conclusion
In this study, we investigated the anti-inflammatory activity of KOG and its mechanism in the inflammatory response of microglia induced by LPS.
KOG inhibited the production of nitrite in LPS-induced BV2 microglia.
KOG inhibited the mRNA expression of iNOS and COX2 in LPS-induced BV2 microglia.
KOG inhibited the mRNA expression of IL-1 β, IL-6 and TNF-α in LPS-induced BV2 microglia.
KOG inhibited the phosphorylation of JNK, p38 and the degradation of Iκ-Bα in LPS-induced BV2 microglia.
The above results show that KOG has an anti-inflammatory activity through the inhibition of JNK and p38 phosphorylation and Iκ-Bα degradation in LPS-induced BV2 microglia (Fig. 6). Therefore, it is thought that KOG can be applied to the development of new medicine related to neuronal inflammation and neurodegenerative diseases. Also, this study could be basic data in clinical indication expansions of KOG.