Yang, Jin, Kwon, Cho, Park, Jung, Moon, Park, Cho, and Ko: Experiment on the effect of Artemisia sieversiana extract on hair loss prevention and cell growth
Original Article
The Journal of Korean Medicine 2022; 43(1): 18-32.
1Department of Korean Internal Medicine, College of Korean Medicine, Gachon University
2Department of Cardiology and Neurology, College of Korean Medicine, Kyung Hee University
3Department of Korean Internal Medicine, Stroke and Neurological Disorders Center, Kyung Hee University Hospital at Gangdong, Kyung Hee University
Correspondence to : Chang-Nam Ko, Department of Korean Internal Medicine, Stroke and Neurological Disorders Center, Kyung Hee University Hospital at Gangdong, 892 Dongnam-ro, Gangdong-gu, 05278 Seoul, Korea Tel : +82 2 440 6215, Fax : +82 2 440 7171, E-mail: kcn202@khu.ac.kr
Received October 5, 2021 Revised January 11, 2022 Accepted February 22, 2022
Abstract
Objectives
This study aimed to examine the safety, effects on proliferation of hair papilla cells, and anti-inflammatory and antioxidant mechanisms of Artemisia sieversiana Ehrh. ex Willd. (AS) extract.
Methods
Safety tests through purity testing, acute toxicity tests, and repeated toxicity tests were performed using AS extract (ASE) which had been dried for over two years. Cell culture and proliferation tests were conducted; VEGF (vascular endothelial growth factor), bFGF (basic fibroblast growth factor), and EGF (epidermal growth factor) and protein expression analyses were performed for mechanistic evaluation; and inhibitory effects of ASE on the RNA expression of testosterone, 5α-reductase, and aromatase was assessed. The anti-inflammatory and antioxidant efficacy of ASE was confirmed by measuring the levels of nitric oxide, inflammatory mediators (TNF-α and PGE2), inflammatory cytokines (IL-1β, IL-6, and IL-8), and chemokine MCP-1.
Results
The safety of ASE was confirmed. The mechanism of cell proliferation in human hair follicle dermal papilla cells involved the promotion of VEGF, bFGF, and EGF expression. ASE decreased mRNA expression of testosterone, 5α-reductase, and aromatase-1 in a concentration-dependent manner. PGE2 and TNF-α production by inflammatory mediators was also significantly decreased in a concentration-dependent manner, and inflammatory cytokine and chemokine expression was inhibited.
Conclusions
ASE is suggested to promote papillary cell growth at the cellular level, to suppress expression of various enzymes involved in hair cycle and cell death, and to inhibit hair loss through anti-androgen, anti-inflammatory, and antioxidant effects.
Artemisia sieversiana Ehrh. ex Willd. (AS), also called Daejaho (大杍蒿), is a medicinal herb belonging to the Asteraceae family. Unlike other medicinal herbs, only the leaves of AS can be used in the first year, and stems and roots can be used in the second year. AS is distributed in some areas of Korea, China, and Japan and has been used for the treatment of bronchitis, tonsillitis, mastitis, dermatitis, hepatitis, and urinary tract inflammation. It consists of terpenoids, lignans, flavonoids, steroids, and alkaloids, all of which have anti-inflammatory, antioxidant, and antibacterial effects.1)
Hair loss, which is increasing year by year, has increased the cost burden for treatment; however, there is no effective preventive method. The only drugs approved by the FDA are minoxidil and propecia, both of which are ineffective in preventing hair loss; therefore, there is a growing interest in developing novel treatments, including herbal medicines. Hair loss is mainly caused by autoimmune mechanisms, neuropsychiatric factors, genetic factors, and hormonal imbalances.2) Androgenic alopecia accounts for most cases of hair loss and is caused by dihydrotestosterone (DHT) converted from testosterone, which atrophies the hair follicles and gradually thins the hair.3)
Experimental studies have been conducted to investigate the effect of herbal medicine extracts, such as Pinelliae rhizoma (半夏) and Arisaema peninsulae (天南星) on hair loss through macroscopic observation or the observation of hair follicles using an optical microscope.4,5) In a recent study, the inhibition of 5α-reductase 1,2 of Sophora flavescens extract,6) hair growth stimulation due to mixed herbal medicines, such as Dangguibohyul-tang gami (當歸補血湯加味)7) and Gamissanghwa-tang (加味雙和湯)8), which inhibited the activity of 5α-reductase type 2 to suppress hair loss and inhibited nitric oxide (NO) production were reported.
This study experimentally verified the efficacy, safety, toxicity, proliferation, and mechanism of dermal papilla cells involved in the hair cycle, anti-androgen 5α-reductase, aromatase enzyme activity, and anti-inflammatory and antioxidant mechanisms of AS extract (ASE).
Research materials and methods
1. Research Materials
Artemisia sieversiana Ehrh. ex Willd. (AS), grown and collected in Gangwon-do, Republic of Korea, was purchased from Modam Global Nature Co. The plants had been cultivated by this company and had been dried for over two years. The whole plants, including roots, stems, and leaves, were boiled at a ratio of 37.2 g per 1 L of water for 2 hours and 250 mL of decoction was prepared.
2. Research Method
2.1. Safety test
This test was conducted to check the safety of AS by collecting the raw materials of AS and conducting a purity test (heavy metal and residual pesticide) (Table 1).
2.2. Acute toxicity and repeated toxicity test
2.2.1. Acute toxicity test
To investigate the toxicity of the ASE and confirm the lethal dose, oral administration of the ASE was performed in male and female Sprague-Dawley (SD) rats. The mortality, general symptoms, and weight change were observed for 14 days after a single oral administration at doses of 0, 500, 1000, and 2000 mg/kg to five male rats and ten female rats per group. After the observation period, an autopsy was performed to observe the visual findings.
2.2.2. Repeated dose toxicity test
Male ICR mice were fasted from the evening prior to the experiment to facilitate drug absorption, and were divided into groups by body weight on the day of the experiment. Subsequently, the ASE was orally administered at a concentration of 2 g/kg per body weight, and abnormal behavior was observed for more than 4 h. The drug was then administered orally for 5 days.
In the case of mortality of a subject, an autopsy was performed immediately, and even if no lethal case occurred, the anomaly was over-anesthetized with ether the subsequent day after drug administration. Additionally, the department visually observed for abnormalities in the organs compared to the control group.
2.3. Cell culture and proliferation test
2.3.1. Cell culture
Human epidermal keratinocytes and human hair follicle dermal papilla cells purchased from Applied Biological Materials (Richmond, British Columbia, Canada) were added to the DMEM culture medium by adding 100 U/ml penicillin-100 U/ml streptomycin and 10% fetal bovine serum (FBS, Invitrogen) and cultured in a 5% CO2 incubator.
2.4. Mechanism study through VEGF, bFGF, EGF, and protein expression analysis
To investigate the mechanism of cell proliferation in human hair follicle dermal papilla cells, VEGF (vascular endothelial growth factor) 50 ng/mL, bFGF (basic fibroblast growth factor) 50 ng/mL, and EGF (epidermal growth factor) 20 ng/mL were respectively added to the cultured cells after dispensing the cultured cells into a 6-well plate and cultured for approximately 90%. After treatment for the first time, treatments of 62.5, 250, and 500 μg/mL of ASE were administered and incubated in a CO2 incubator for 48 h.
After cultivation, the expression levels of VEGF, bFGF, and EGF were confirmed by ELISA in the culture supernatant, and protein expression was confirmed by western blot assay by extracting proteins from the cells.
2.5. Analysis of RNA expression inhibition of testosterone, reductase, and aromatase of ASE
To confirm the inhibition of testosterone, reductase, and aromatase-1 activity in human hair follicle dermal papilla cells, the cells were dispensed into 6 well plates when 90% of the cells were cultured. Subsequently, a treatment with EGF (20 ng/mL) was first performed for 2 h, and then treatment with ASE (62.5 μg/mL, 250 μg/mL, and 500 μ g/mL) was performed and incubated in a CO2 incubator for 48 h. After culturing, the total RNA was extracted from the cells and RNA expression was confirmed by RT-PCR.
2.6. Anti-inflammatory, antioxidant, and mechanism studies in human keratinocytes with ASE
The cells were dispensed in a 96-well plate, such that the number of cells was 1×105 per well; the cells were incubated at 90% confluence in a CO2 incubator. After washing each well twice with DMEM to remove non-adherent cells, the ASE was diluted by 0–500 μg/mL concentration. After adding 100 μL of ASE to each well, 1 μg/mL of LPS and 1 ng/mL of IFN-γ were added to stimulate the cells, and then cultured in a CO2 incubator for 72 h. This was analyzed using the MTT assay and was analyzed at 48 h and 92 h.
2.6.2. Measurement of nitric oxide (NO) production
The cells were treated with 62.5 μg/mL and 250 μg/mL of ASE under the same conditions as the cell proliferation inhibition experiment, treated with LPS 1 μg/mL and IFN-γ 1 ng/mL after 30 min, and cultured in a CO2 incubator for 72 h. Subsequently, the amount of NO in the culture supernatant was measured using the Griess reagent. Notably, 100μL of culture medium and 100 μL of Griess reagent (1% sulfanilamide + 0.2% N-naphthyl-ethylenediamine 2HCl + 2.5% H3PO4) were mixed and dispensed into a new 96-well plate, and the absorbance was measured within 10 min at 570 nm. A standard calibration curve was prepared using NaNO2 to determine the amount of NO produced by the sample.
2.6.3. Measurement of inflammatory mediators, TNF-α and PGE2
After the macrophages were dispensed into a number of 106 cells per well in a 24-well plate, 100 μg/mL of RAH13 drug and 100 nM of celecoxib as a positive control were treated, and after 30 min, the cells were treated with 1 μg/mL LPS, and 1 ng/mL IFN-γ. After incubation in a CO2 incubator for 24 h, the supernatant was collected by centrifugation at 2000 rpm for 5 min and used for cytokine measurement. The amount of TNF-α (BD Pharmingen Inc., USA) and PGE2 (R&D Systems, USA) in the collected supernatant were measured by an enzyme immunoassay using a commercially available kit.
2.6.4. Measurement of inflammatory cytokines (IL-1β, IL-6 and IL-8) and chemokine MCP-1
Keratinocytes were treated with 1 μg/mL of LPS and 1 ng/mL of IFN-γ and cultured in a CO2 incubator for 24 h, followed by treatment with 62.5 μg/mL and 250 μg/mL of ASE and incubation for 48 h. After incubation, the plate was centrifuged at 2000 rpm for 5 min to collect the culture supernatant. The levels of IL-1β, IL-6, IL-8, and MCP-1 in the culture medium were measured by enzyme immunoassay using a commercially available kit (BD Pharmingen Inc., USA).
2.6.5. Analysis of signaling mechanism of anti-inflammatory and antioxidant efficacy of ASE
The keratinocytes were treated with 1 μg/mL of LPS and 1 ng/mL of IFN-γ and incubated in a CO2 incubator for 24 h, treated with 62.5 μg/mL and 250 μg/mL of ASE, and incubated again for 18 h. After incubation, the proteins were collected from the cells. After protein quantification, 20 μg was electrophoresed on a 4–12% acrylamide gel, and the proteins were transferred to nitrocellulose paper. After diluting p-ERK, p-p38, p-JNK, and Iκ Bα antibodies at 1:1000, they were allowed to react for 24 h. After washing three times in PBST solution, a secondary antibody was attached, and luminescence was detected using chemiluminescence.
7. Statistics processing
All values are shown as means ± standard deviation. An independent samples t-test was used, and statistical significance is reported at p < 0.05.
Results
1. Safety test results
As a result of confirming the safety by performing a pesticide residue test and heavy metal test by purity testing, the residual amount of pesticide was determined to be less than 0.1 ppm of the total DDP, less than 0.01 ppm of dieldrin, less than 0.2 ppm of total BHC, less than 0.01 ppm of aldrin, and less than 0.01 ppm of endrin. The safety test confirmed that there were no abnormalities in its use. The heavy metal test showed that the level of arsenic (As) was less than 3 ppm, level of cadmium (Cd) 0.3 ppm or less, and level of mercury (Hg) was 0.2 ppm or less. The safety was confirmed as no abnormality in the use of AS (Table 1).
2. Results of acute toxicity tests and repeated toxicity tests
2.1. Results of acute toxicity test
The mortality, general symptoms, and body weight change were observed for 14 days after a single oral administration of AS at doses of 0, 500, 1000, and 2000 mg/kg for 10 males and females in each group. No specific changes were observed in the general symptom weight changes or autopsy findings.
2.2. Results of repeated dose toxicity test
In the high-dose repeated-dose toxicity test of 5 g/kg of ASE, no mortality of the subjects occurred, and the side effects or abnormalities of organs were not observed with the naked eye during administration. It was found that the ASE did not induce repeated dose toxicity after oral administration. After the end of the observation period, an autopsy was performed, and no gross organ abnormalities were observed.
3. Results of cell proliferation and the mechanism of ASE
3.1. Cell proliferation results of ASE
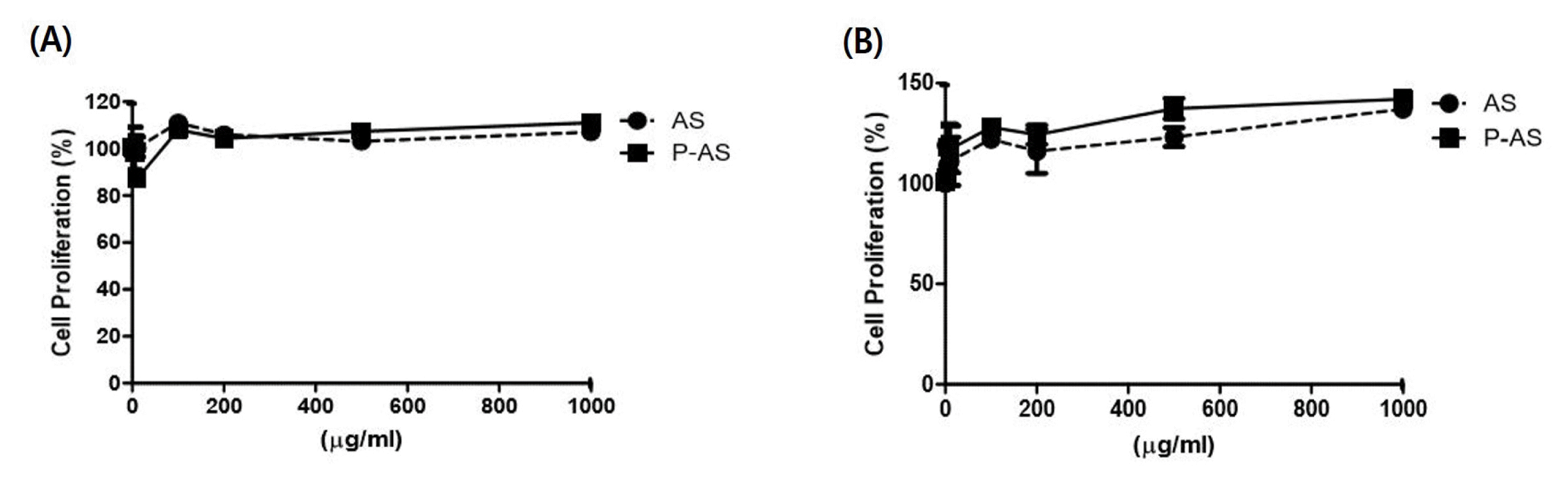
In human epithelial keratinocytes, there was no significant cell proliferation in the ASE and AS added (P-AS) (Fig. 1A). However, in human hair follicle dermal papilla cells, the ASE and AS added prescription (P-AS) increased the cell proliferation by 23% to 55% at concentrations of 250 μg/mL, 500 μg/mL, and 1000 μg/mL (Fig. 1B).
3.2. Mechanisms of cell proliferation
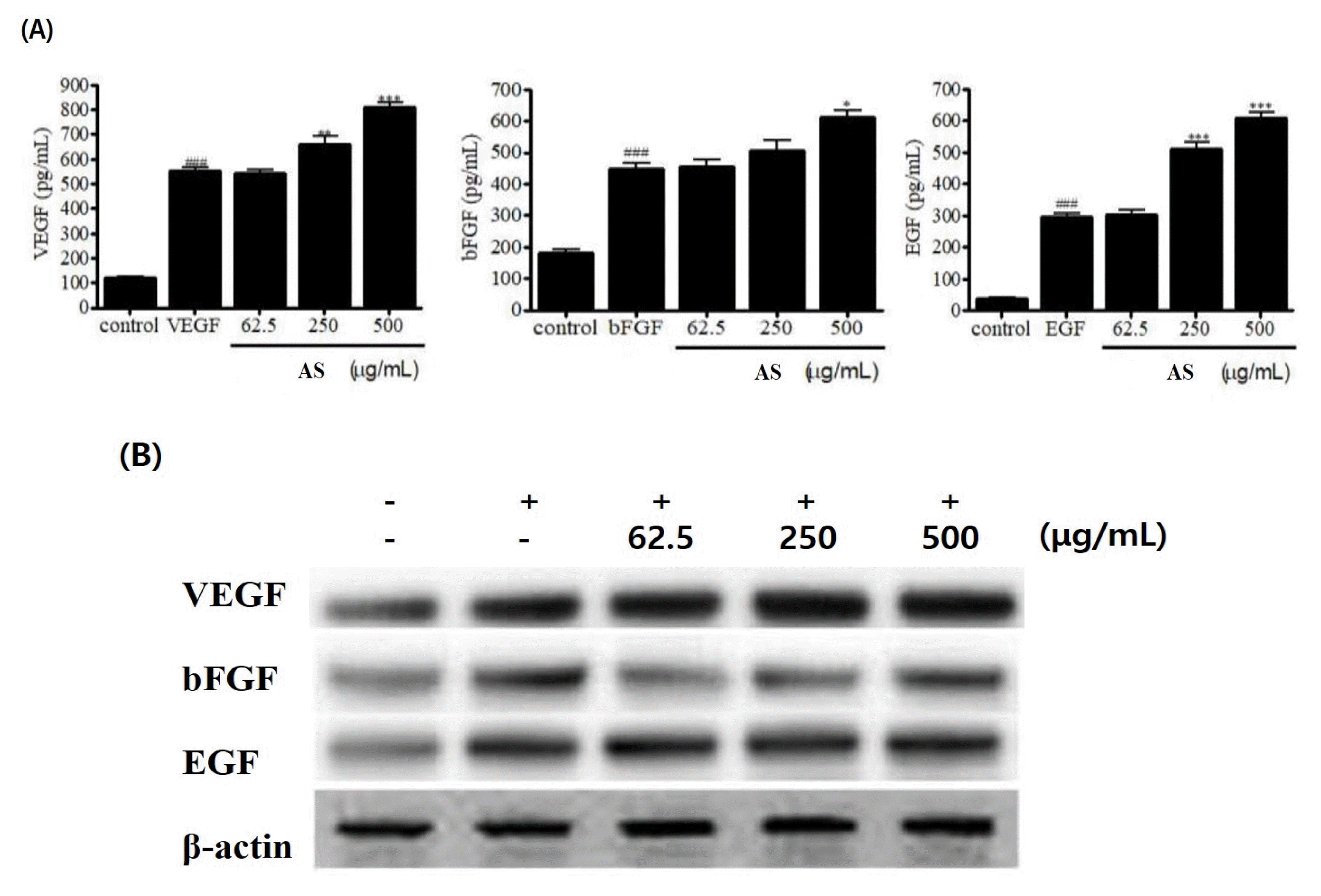
The ASE significantly increased the expression of VEGF at 250 and 500 μg/mL; additionally, the expression of bFGF was significantly increased at 500 μg/mL. The EGF expression was significantly increased at 250 μg/mL and 500 μg/mL; it was confirmed that the cell proliferation in human hair follicle dermal papilla cells was due to the promotion of VEGF, bFGF, and EGF expression (Fig. 2).
4. Results on mRNA activity of testosterone, reductase, and aromatase of ASE
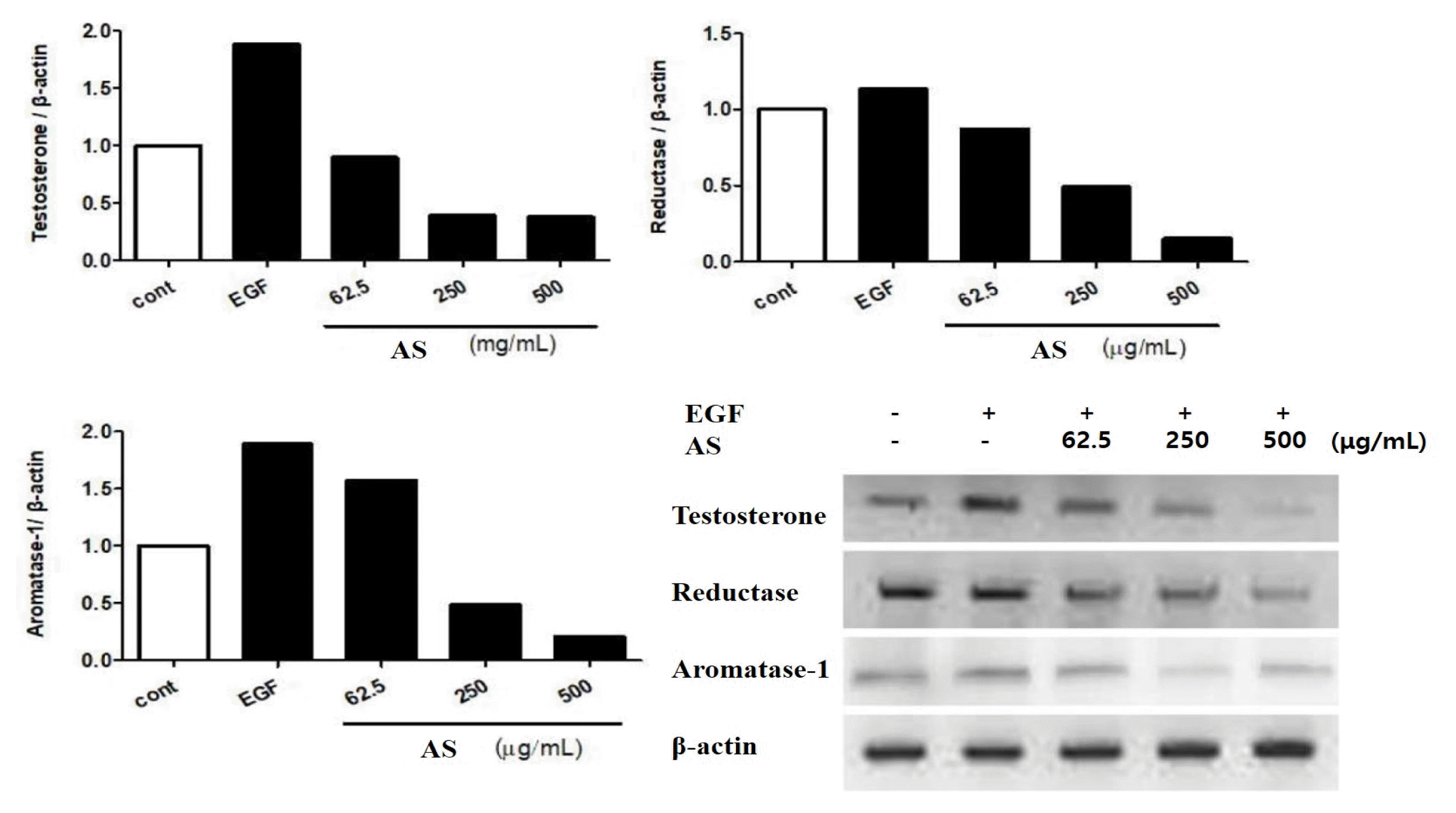
The ASE decreased the mRNA expression of testosterone, reductase, and aromatase-1 at 62.5, 250, and 500 μg/mL, respectively (Fig. 3).
5. Results of anti-inflammatory, anti-oxidative, and mechanism studies in human keratinocytes of ASE
5.1. Results of keratinocyte proliferation inhibition test
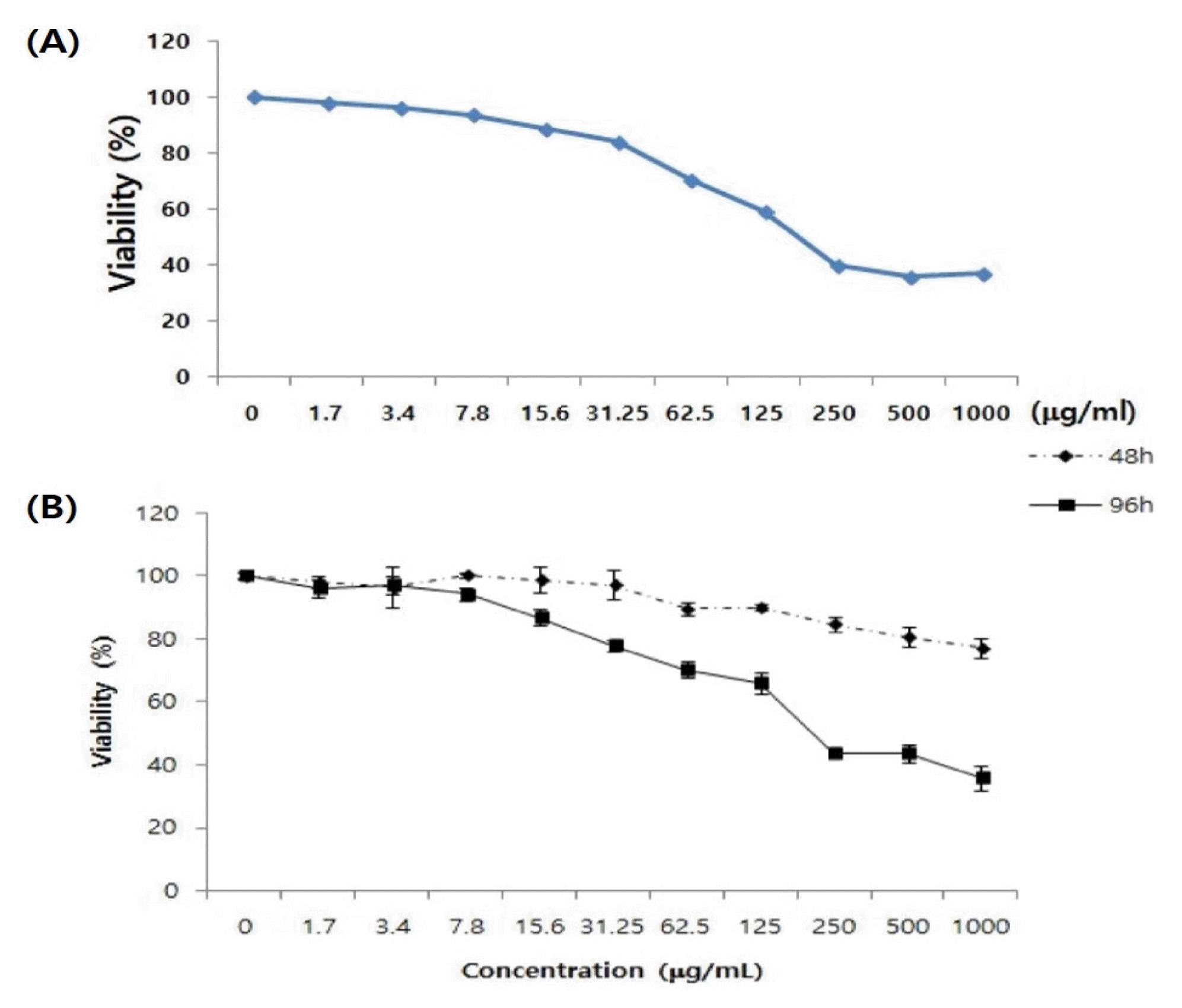
In inflammatory keratinocytes, it was confirmed that 30% of the cell proliferation was inhibited at 62.5 μg/mL of ASE and that cell proliferation was inhibited by 69.1% at a concentration of 250 μ g/mL (Fig. 4A). In addition, it was confirmed that an excellent inhibitory effect was observed up to 92 h in the hourly growth inhibition experiment (Fig. 4B).
5.2. NO production inhibitory effect
The amount of NO that induces inflammation by promoting the secretion of inflammatory cytokines showed significant inhibitory effects of 14.3% and 38.8%, depending on the concentration of the ASE at 62.5 μg/mL and 250 μg/mL (Table 2).
5.3. Results of inflammatory mediators (TNF-α and PGE2) measurement
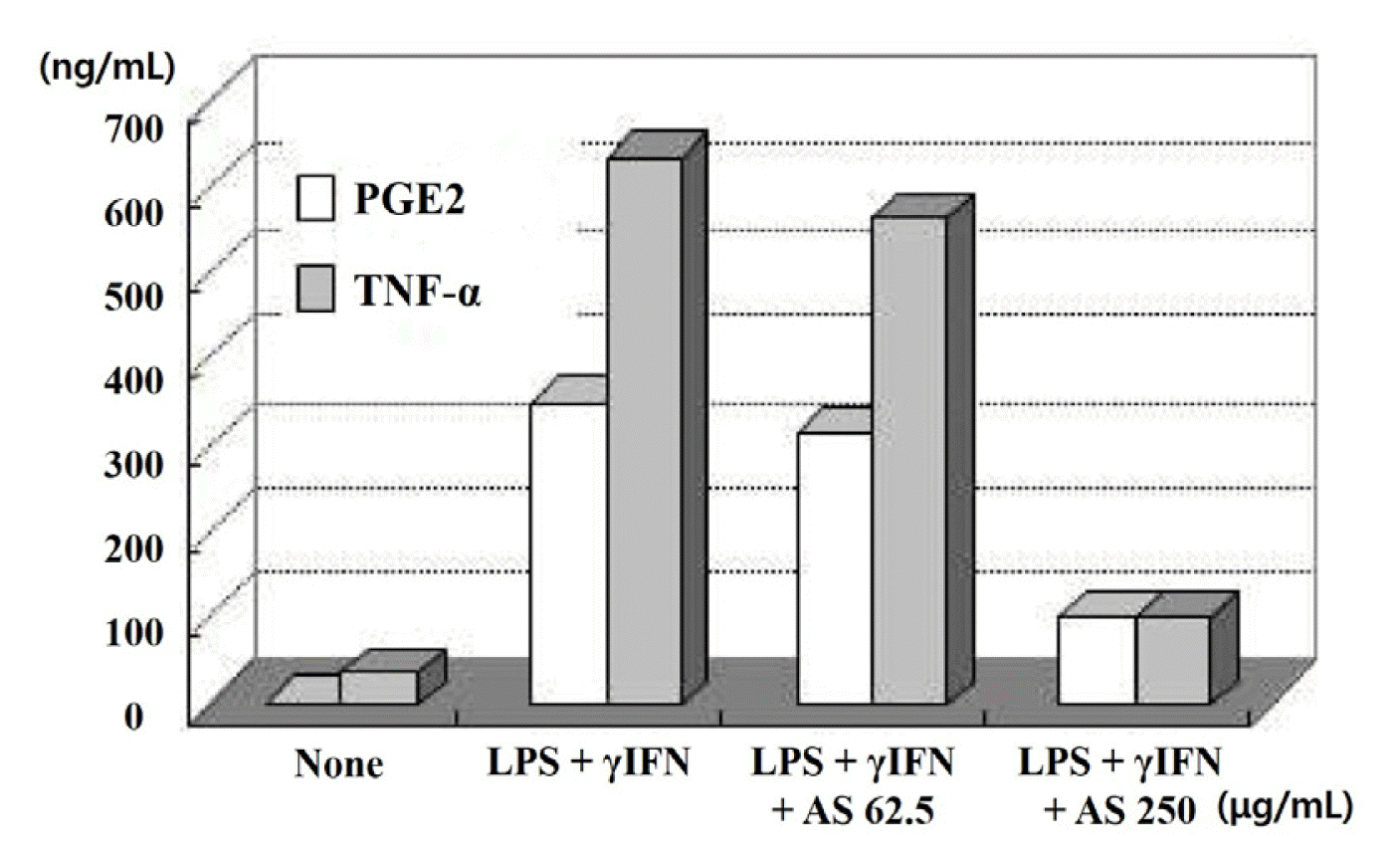
PGE2 and TNF-α production was reduced by approximately 10% and 17%, respectively, at 62.5 μg/mL ASE. At 250 μg/mL, PGE2 and TNF-α levels were significantly reduced to approximately 35% and 77%, respectively (Fig. 5).
5.4. Results of inflammatory cytokines (IL-1 β, IL-6, and IL-8) and chemokine MCP-1 measurement
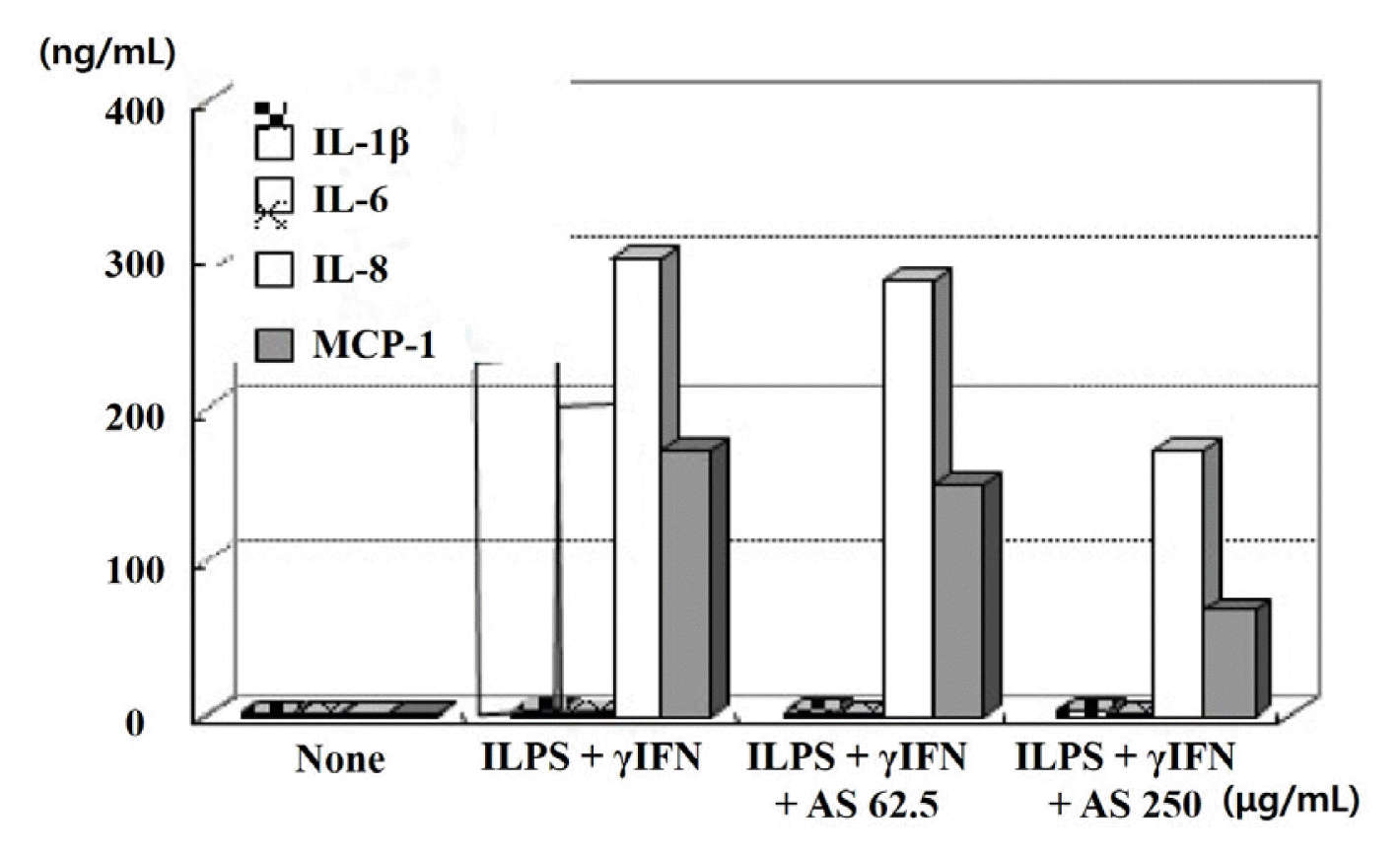
The ASE inhibited IL-1β and IL-6, which are inflammatory cytokines, by almost 100%, not only at low concentrations (62.5 μg/mL), but also showed significant reduction in the expression of IL-8 and MCP-1 (40.3% and 69%), respectively at 250 μg/mL (Fig. 6).
5.5. Mechanism of signaling anti-inflammatory and antioxidant efficacy of ASE
The phosphorylation of p-p38 and p-JNK decreased in a concentration-dependent manner, and IκBα tended to decrease only with administration of ASE of concentration of 62.5 μg/mL. As a result, the anti-inflammatory and antioxidant efficacy in inflammatory keratinocytes is thought to be due to the inhibition of p-p38, p-JNK, and IκBα signaling (Fig 7).
Discussion
Artemisia sieversiana Ehrh. ex Willd. (AS) is a herbaceous plant belonging to the family Mugwort and has a stronger scent than thrush, which is used to treat malaria. In Korea, it can be seen occasionally in the southern regions and grows in the middle or north. In China, it is found mainly in the northeastern or northern areas. It is a medicinal herb that is used to treat hemoptysis, asthma, jaundice, diarrhea, sore throat, urinary tract inflammation, soreness, numbness, pain, and trauma due to hemorrhage. It contains sieversinin, a sesquiterpene found in the leaves and stems, and absinthin. Dried AS contains alkaloids (0.12–0.2%), pyrocatechol tannin (0.499%), flavonoid (0.831%), lactone, rutin, and isoquercetin.9)
As an experimental study on pharmacological action, sesquiterpene and y-lactone separated from AS have an in vitro inhibitory effect against Staphylococcus aureus, E. coli, etc.9) Rutin showed anti-cancer effects by preventing G2/M phase induction of the cell cycle and regulating the expression of genes related to apoptosis.10) It was determined that absinthin inhibited the accumulation of inflammatory cells in lung tissue.11) In addition, beta-sitosterol has antibacterial effects on Salmonella typhi, Corynebacterium diphtheriae, etc., in antibacterial tests.12)
Leaves contain secondary metabolites, such as sieversinin, which has antibacterial properties.13) Achillin and leucodin showed potent inhibitory activity when beta-hexosaminidase was released from RBL-2H3 cells. Quercetin is the most notable anti-allergic component and has been found to have anti-allergic efficacy as an antagonist of LTB4.14) Sesamin did not show antioxidant efficacy in in vitro experiments; however, in vivo experiments showed that it has a particularly protective effect on the antioxidant activity of the liver. This suggests that sesamin may be an effective substance to prevent oxidative stress in the liver.15) In addition, chrysoeriol reduced the ROS production in PMN cells, producing large amounts of ROS during oxidative stress.16) As such, in previous studies, experimental studies on single components of AS have been reported; however, studies using ASE are insufficient, and it is thought that the anti-inflammatory, anti-androgen, and antioxidant effects of ASE can be experimentally verified to be effective in hair loss. The purpose of this study was to clarify the ability to promote the proliferation of white papillae dermal papilla cells and their mechanism, and to quantitatively verify the efficacy of anti-androgen 5α-reductase, aromatase enzyme activity inhibition, anti-inflammatory, and antioxidant mechanisms.
To confirm safety, the results of the pesticide residue and heavy metal tests confirmed that there was no abnormality when using the ASE. Looking at the results of the acute toxicity and repeated-dose toxicity tests to confirm the results of the toxicity test, no specific changes were observed in the general symptoms, weight change, and autopsy findings in the acute toxicity test, and no mortality was observed even at high concentrations of the extract. In the repeated dose toxicity test results, no mortality of the subjects occurred, repeated dose toxicity was not expressed by oral administration, and no gross organ abnormalities were observed in the autopsy results. This confirmed that the safety was secured even in acute toxicity and repeated dose toxicity through oral administration of the ASE.
The hair follicle dermal papilla cells play an important role in the cycle regulation of hair follicles, and in the case of male pattern hair loss, the male hormone receptors are mainly distributed in the hair follicles of the hair follicles. Therefore, male hormones act on the hair papillae to secrete growth inducer of hair. Hair loss occurs by the inhibiting the growth of dermal papilla cells.3,17) Based on the results of cell proliferation and mechanism studies in this report, there was no significant cell proliferation in human epithelial keratinocytes in the AS and the prescription with the addition of P-AS in the cell proliferation results. However, treatment with AS and the addition of P-AS increased the cell proliferation by 23–55% in a concentration-dependent manner in human hair follicle dermal papilla cells. These results are thought to promote the hair growth through the proliferation of dermal papilla cells.
Based on this, the expression of VEGF and EGF was significantly increased in a concentration-dependent manner, and the expression of bFGF was also significantly increased at a high concentration in the cell proliferation mechanism study. It was found that the mechanism of cell proliferation in human hair follicle dermal papilla cells is due to the promotion of VEGF, bFGF, and EGF expression. It is thought that these results promote the proliferation of dermal papilla cells and thus may be involved in hair follicle cell growth.
Currently, clinically applied alopecia treatments can be viewed in two major ways: first, such treatments promote the growth and differentiation of follicular epithelial cells, shorten the telogen, and promote anagen entry, so that the growth phase is long.18) The mechanisms to sustain and inhibit 5 α-reductase prevent the conversion of testosterone to DHT, which is known to be the leading cause of hair loss.19)
Testosterone reaches the hair follicles and changes into a more potent form of DHT by 5α -reductase.20) 5α-reductase exists in two forms, differing in biochemical and pharmacological properties, distribution along organs, and hair loss. Testosterone introduced into the hair follicle from the hair loss site is converted to DHT by 5α -reductase, thereby inhibiting the hair growth. The converted DHT binds to the male hormone receptor; additionally, several male hormone receptors are distributed in the hair loss region, and the male hormone receptor is mainly distributed in the hair follicles. Therefore, if there are several 5α-reductases or many receptors, the probability of progressing to hair loss increases. As a result of examining whether the mRNA expression of testosterone, 5α-reductase, and aromatase-1 has an inhibitory effect, the ASE decreased the mRNA expression of testosterone, 5α-reductase, and aromatase-1 in a concentration-dependent manner. These results confirmed that the ASE inhibits 5α -reductase conversion and testosterone production, which converts testosterone to dihydrotestosterone, suggesting that it can be utilized for hair loss.
In order to investigate the anti-inflammatory and antioxidant efficacy and mechanism of action in human keratinocytes, an MTT assay was performed. In the results analyzed by time, the ASE inhibited the cell proliferation in inflammatory keratinocytes by 30%–69.1%, and proliferated over time. The inhibition test results showed that the inhibitory effect lasted up to 92 h. Through this result, it was found that upon removing the inflammation by inhibiting cell proliferation lasting up to 92 h, the inflammatory effect lasted approximately 3–4 days.
Inflammation is a defense mechanism developed to localize and repair damage to living tissue when it is damaged. It involves a number of complex steps involving blood vessels, nerves, bodily fluids, inflammatory cells, etc., and is ultimately essential for survival to eliminate, dilute, and block causative factors.21) Inflammatory responses are induced by infection or trauma, physical and chemical damage, or autoimmunity. In this process, various triggers, such as pro-inflammatory mediators are involved.22)
At the beginning of the inflammatory response, macrophages are activated to induce the expression of pro-inflammatory cytokines, such as TNF-α, and increase the levels of inflammatory mediators, such as NO and PGE2. At this time, the levels of inflammatory mediators, such as iNOS, cyclooxygenase-2 (COX-2), TNF-α, IL-1, and IL-6 increase through cell signaling. Additionally, it is observed that excessive amounts of pro-inflammatory cytokines or NO also play an important role,23) and excessive production of NO can cause tissue damage in chronic inflammatory diseases.24,25) In addition, the inhibition of pro-inflammatory cytokines and iNOS gene expression in the inflammatory cells is believed to advantageously inhibit the inflammatory response.23)
LPS exists in the outer layer of the cell wall of gram-negative bacteria and acts as an endotoxin that can induce inflammation. Additionally, macrophages produce NO by the induction of iNOS, which can be induced by cytokines, such as IL-1β and TNF-α.26) LPS binds to macrophages CD14/TLR4/MD2 and induces the phosphorylation of IL-1RI-associated protein kinases (IRAK) and activation of TNFR-associated factor (TRAF) 6 through MyD88 and phosphorylation of I-κB. As it produces various cytokines, it is frequently used for inflammation research using macrophages27,28) and binds to the CD14/TLR4/MD2 complex, so that the inflammatory cells activate inflammatory mediator cytokines and secretion of inflammatory mediators, such as NO and eicosanoids.29) The amount of NO that induces inflammation by promoting the secretion of inflammatory cytokines showed a significant inhibitory effect of 14.3% and 38.8%, depending on the concentration of the ASE. It is overproduced in the inflammatory site by the activity of TNF-α, a pro-inflammatory cytokine, and COX-2, which are closely related to the progression of inflammation. As a result of measuring the amount of PGE2 inducing inflammation and pain, the PGE2 and TNF-α production decreased by approximately 10% and 17%, respectively, at low concentrations of the white tiger extract, respectively, and the PGE2 and TNF-α production at high concentrations was approximately 35%, which was significantly reduced to 77%. The results of the measurement of inflammatory cytokines (IL-1β, IL-6, IL-8) and chemokine MCP-1, the ASE inhibited inflammatory cytokines IL-1β and IL-6 by almost 100% at low concentrations. Additionally, the IL-8 and MCP-1 expression decreased at high concentrations (40.3% and 69%, respectively). NO is involved in immune regulation, vasodilation, and neurotransmission. If it is continuously produced in the body or maintained at high pulsation, it may cause excessive inflammatory reactions, which can lead to pathological changes.30) The inflammatory mediators, such as TNF-α, IL-6, IL-1, IL-12, IL-8, and PGE2 play an important role in the initial inflammatory response.31) Therefore, it was confirmed that the ASE prevents an excessive production of NO, prevents cell damage, reduces the amount of production, reduces the cell damage caused by NO, and decreases the production of pro-inflammatory factors PEG2 and TNF-α, thereby exhibiting anti-inflammatory efficacy. In addition, the phosphorylation of p-p38 and p-JNK decreased in a concentration-dependent manner, and IκBα tended to decrease only at 62.5 μg/mL. As a result, the anti-inflammatory and antioxidant efficacy in the inflammatory keratinocytes is thought to be due to the inhibition of p-p38, p-JNK, and IκBα signaling.
From the above, it is presumed that the ASE promotes the growth of papillary cells at the cellular level, suppresses the expression of various enzymes involved in the hair cycle and cell death, and inhibits hair loss through anti-androgen, anti-inflammatory, and antioxidant effects. In the future, we hope that various efficacy studies will be conducted based on the ingredients of AS.
Conclusions
In the acute and repeated dose toxicity experiments, the toxicity of the ASE was not expressed by oral administration.
The ASE and P-AS were observed to proliferate hair papilla cells.
The mRNA expression of testosterone, 5α-reductase, and aromatase-1 was decreased in the ASE.
The ASE inhibited the cell proliferation (range, 30–69.1%) in a concentration-dependent manner in inflammatory human keratinocytes; additionally, the inflammation inhibitory effect lasted up to 92 h.
Nitric oxide production induced inflammation by promoting the secretion of inflammatory cytokines and showed a significant inhibitory effect depending on the concentration of the ASE.
PGE2 and TNF-α production of the inflammatory mediators were significantly decreased in a concentration-dependent manner.
The ASE inhibited the expression of inflammatory cytokines (IL-1β, IL-6, IL-8) and chemokine MCP-1.
The anti-inflammatory and antioxidant efficacy mechanisms are presumed to be due to the inhibition of p-p38, p-JNK, and IκBα signaling.
Fig. 1
Effect of cell proliferation
A: Human epithelial keratinocytes, B: Hair follicle dermal papilla cell
AS: Artemisia sieversiana Ehrh. Ex Willd., P-AS: AS-added prescription
Fig. 2
Confirmation of VEGF, bFGF, and EGF expression before growth propagation of AS extract; (A) ELISA assay (B) Western blot assay
AS: Artemisia sieversiana Ehrh. Ex Willd.
Fig. 3
Quantitative analysis of testosterone, reductase, and aromatase activity inhibitory activity of AS extract at mRNA level
AS: Artemisia sieversiana Ehrh. Ex Willd.
Fig. 4
Inhibitory effect of white tiger cell proliferation on keratinocytes; (A) MTT assay (72 h analysis) (B) MTT assay (48 h and 92 h analysis)
Fig. 5
Effects of AS extract on PGE2 and TNF-α in LPS + γIFN stimulated keratinocytes
AS: Artemisia sieversiana Ehrh. Ex Willd.
PGE2 and TNF-α production was reduced by approximately 10% and 17%, respectively, at 62.5 μg/mL AS extract. At 250 μg/mL, PGE2 and TNF-α levels were significantly reduced to approximately 35% and 77%, respectively
Fig. 6
Effects of AS extract on cytokines expression in LPS+γIFN stimulated keratinocytes
AS: Artemisia sieversiana Ehrh. Ex Willd.
The AS extract inhibited IL-1β and IL-6, by almost 100%, not only at low concentrations (62.5 μg/mL), but also showed significant reduction in the expression of IL-8 and MCP-1 (40.3% and 69%), respectively at 250 μg/mL.
Fig. 7
AS extract inhibits the signal pathway of IκBα, JNK, and p38 MAPK pathway
1. Yong SH, Park DJ, Yang WH, Seol YW, Choi EJ, Jeong MJ, et al. 2019; Cryopreservation of sievers wormwood (Artemisia sieversiana Ehrh. Ex Willd.) seeds by vitrification and encapsulation. Forest Science and Technology. 15:4. 180–186.
https://doi.org/10.1080/21580103.2019.1658647
2. Madani S, Shapiro J. 2000; Alopecia areata update. J Am Acad Dermatol. 42:4. 549–566. quiz 567–570. https://doi.org/10.1067/mjd.2000.103909
3. Randall VA, Hibberts NA, Thornton MJ, Hamada K, Merrick AE, Kato S, et al. 2000; The hair follicle: a paradoxical androgen target organ. Horm Res. 54:5–6. 243–250.
https://doi.org/10.1159/000053266
4. Jeong IK, Jo HY, Kim TH, Kim NS, Jeong HS, Lee CH. 2009; Experimental Studies on the Hair Growth Activity of Extracts of Pinelliae Rhizoma in Spontaneous Alopecia Model and Normal C57BL/ 6N Mice. Korean J Orient Physiol Pathol. 23:1. 84–92.
5. Kwon KS, Lee MW, Jeong IK, Jeong HS, Song BY, Song JM, et al. 2009; Experimental studies on the hair growth activity of fractions and extract of arisaematis rhizoma in C57B/6N mice. Korean J Orient Physiol Pathol. 23:3. 619–630.
6. Roh HC, Roh SS. 2002; Studies on the effect of Sophora flavescens extract on the hair growth stimulation and acne inhibition. J Korean Med Ophthalmol Otolaryngol Dermatol. 15:1. 96–126.
7. Kim PS, Kim HT, Roh SS, Hwang CY. 2004; Effect of DanGuiBoHyulTangGami-Bang on the Alopecia and Hair Growth Stimulation. J Korean Med Ophthalmol Otolaryngol Dermatol. 17:3. 38–60.
8. Yun JH, Kim NK, Lim KS, Roh SS, Hwang CY. 2004; Study on the effect of Gamissanghwa-tang and each medicinal plant extract for the hair growth of the mice using in vivo and in vitro test. Korean J Orient Physiol Pathol. 18:2. 561–570.
9. Liu S-J, Liao Z-X, Tang Z-S, Cui C-L, Liu H-B, Liang Y-N, et al. 2017; Phytochemicals and biological activities of Artemisia sieversiana. Phytochemistry Reviews. 16:3. 441–460.
https://doi.org/10.1007/s11101-016-9475-z
10. Chen H, Miao Q, Geng M, Liu J, Hu Y, Tian L, et al. 2013; Anti-Tumor Effect of Rutin on Human Neuroblastoma Cell Lines through Inducing G2/M Cell Cycle Arrest and Promoting Apoptosis. ScientificWorldJournal. 269165
https://doi.org/10.1155/2013/269165
11. Guo N, Xu Y, Cao Z. 2015; Absinthin attenuates LPS-induced ALI through MIP-1α-mediated inflammatory cell infiltration. Exp Lung Res. 41:9. 514–524.
https://doi.org/10.3109/01902148.2015.1093566
12. Kiprono PC, Kaberia F, Keriko JM, Karanja JN. 2000; The in vitro anti-fungal and anti-bacterial activities of beta-sitosterol from Senecio lyratus (Asteraceae). Z Naturforsch C J Biosci. 55:5–6. 485–488.
https://doi.org/10.1515/znc-2000-5-629
13. Kumar J, Mishra GP, Naik PK, Murkute AA, Srivastava RB. 2011; Genomic DNA isolation from Artemisia species grown in cold desert high altitude of India. Afr J Biotechnol. 10:37. 7303–7307.
https://doi.org/10.5897/AJB10.2031
14. Serra MF, Diaz BL, Barreto EO, Pereira AP, Lima MC, Barbosa-Filho JM, et al. 1997; Anti-allergic properties of the natural PAF antagonist yangambin. Planta Med. 63:3. 207–212.
https://doi.org/10.1055/s-2006-957654
15. Nakai M, Harada M, Nakahara K, Akimoto K, Shibata H, Miki W, et al. 2003; Novel antioxidative metabolites in rat liver with ingested sesamin. J Agric Food Chem. 51:6. 1666–1670.
https://doi.org/10.1021/jf0258961
16. Kang J, Xie C, Li Z, Nagarajan S, Schauss AG, Wu T, et al. 2011; Flavonoids from acai (Euterpe oleracea Mart.) pulp and their antioxidant and anti-inflammatory activities. Food Chem. 128:1. 152–157.
https://doi.org/10.1016/j.foodchem.2011.03.011
17. Mounsey AL, Reed SW. 2009; Diagnosing and treating hair loss. Am Fam Physician. 80:4. 356–362.
18. Hagemann T, Schlütter-Böhmer B, Allam JP, Bieber T, Novak N. 2005; Positive lymphocyte transformation test in a patient with allergic contact dermatitis of the scalp after short-term use of topical minoxidil solution. Contact Dermatitis. 53:1. 53–55.
https://doi.org/10.1111/j.0105-1873.2005.00456b.x
19. Diani AR, Mulholland MJ, Shull KL, Kubicek MF, Johnson GA, Schostarez HJ, et al. 1992; Hair growth effects of oral administration of finasteride, a steroid 5 alpha-reductase inhibitor, alone and in combination with topical minoxidil in the balding stumptail macaque. J Clin Endocrinol Metab. 74:2. 345–350.
https://doi.org/10.1210/jcem.74.2.1309834
20. Rathnayake D, Sinclair R. 2010; Male androgenetic alopecia. Expert Opin Pharmacother. 11:8. 1295–1304.
https://doi.org/10.1517/14656561003752730
21. Korean Society of Pathologists. 1998. Textbook of Pathology. 3rd ed. Seoul: Komoonsa.
22. Kumar Vinay, Abbas Abul K, Fausto Nelson, Aster Jon. 2011. Robbins & Cotran Pathologic Basis of Disease. 8th ed. Philadelphia: Saunders.
23. Southan GJ, Szabo C. 1996; Selective pharmacological inhibition of distinct nitric oxide synthase isoforms. Biochem Pharmacol. 51:4. 383–394.
https://doi.org/10.1016/0006-2952(95)02099-3
24. Chawla A, Nguyen KD, Goh YP. 2011; Macrophage-mediated inflammation in metabolic disease. Nat Rev Immunol. 11:11. 738–749.
https://doi.org/10.1038/nri3071
25. Aktan F. 2004; iNOS-mediated nitric oxide production and its regulation. Life Sci. 75:6. 639–653.
https://doi.org/10.1016/j.lfs.2003.10.042
26. Laskin DL, Pendino KJ. 1995; Macrophages and inflammatory mediators in tissue injury. Annu Rev Pharmacol Toxicol. 35:655–677.
https://doi.org/10.1146/annurev.pa.35.040195.003255
29. Kim YW, Zhao RJ, Park SJ, Lee JR, Cho IJ, Yang CH, et al. 2008; Anti-inflammatory effects of liquiritigenin as a consequence of the inhibition of NF-kappaB-dependent iNOS and proinflammatory cytokines production. Br J Pharmacol. 154:1. 165–173.
https://doi.org/10.1038/bjp.2008.79
30. Wink DA, Mitchell JB. 1998; Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic Biol Med. 25:4–5. 434–456.
https://doi.org/10.1016/s0891-5849(98)00092-6
31. Janeway CA, Travers P, Walport M, Shlomchik MJ. 2001. Immunobiology: The Immune System in Health and Disease. 5th ed. New York: Garland Science.